Последствия антропогенного влияния на состав крови цыплят
реклама

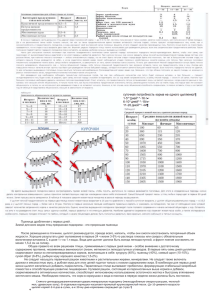
БИОЛОГИЧЕСКИЕ НАУКИ Последствия антропогенного влияния на состав крови цыплятбройлеров А.И. Вишняков, к.б.н., доцент, Оренбургский ГУ; А.А. Торшков, к.б.н., доцент, Оренбургский ГАУ С развитием интенсивных технологий выра щивания и откорма сельскохозяйственной пти цы большое внимание уделяется её полноцен ному питанию, причем не только наличию пи тательных веществ в рационе, но и обращается особое внимание на содержание минеральных веществ и витаминов, а также других биологи чески активных веществ. Такой комплексный подход требует глубоких научных исследова ний системы их применения, совместимости, влияния на состояние обменных процессов в организме, продуктивность и качество про дукции. Известно, что состояние системы красной крови позволяет дать объективную оценку фи зиологического статуса организма в целом [1]. Целью наших исследований было выявить изме нения морфологического состава крови при сни жении в рационе цыплятбройлеров содержания минеральных веществ и обменной энергии. Для этого были созданы три группы цыплят бройлеров по принципу аналогов (кросс, воз раст, живая масса). Из них одна контрольная, получающая основной рацион, и две опытные: первая получала «аминеральный» рацион, а вто рая – рацион с содержанием обменной энергии, переваримого протеина и минеральновитамин ного комплекса на уровне нижней границы нор мы. При расчетах рационов использовали дан ные потребности птицы в питательных веще ствах и микроэлементах [2]. Для изучения мор фологического состава крови убой цыплят про водился в возрасте 10, 20, 30, 40 и 50 суток в количестве по 5 голов. Исследования количества гемоглобина в кро ви цыплят показали, что в возрасте десяти суток его концентрация составляет в среднем 87,3 г/л. В контрольной группе этот показатель к концу исследуемого периода увеличивается до макси мального уровня, который составляет 140,7 г/л. В первой опытной группе максимальная кон центрация гемоглобина зафиксирована в возра сте 40 суток и составляет 135,7 г/л. Во второй опытной группе концентрация гемоглобина ко леблется в пределах от 84,7 г/л (в 20 суток) до 125,3 г/л (в 50 суток). При этом в опытных груп пах содержание гемоглобина в различных возра стных группах на 5,51–21,11% меньше, чем в контрольной, за исключением 40суточного воз раста, когда этот показатель в контрольной груп пе уступает опытным: 14,75–33,43%. Возрастная динамика количества эритроци тов в крови цыплят, в общем, отражает таковую концентрации гемоглобина. В возрасте десяти суток постинкубационного периода количество эритроцитов составляет 1,717.1012/л. В конт рольной группе с возрастом их количество уве личивается и к 50суточному возрасту достигает своего максимума – 3,913.1012/л. У цыплят, содержащихся на «аминеральном» рационе, эти клетки максимального количества в единице объёма крови достигли в возрасте 40 суток (3,860.1012/л). Во второй опытной группе мак симальный показатель, равный 3,913.1012/л, отмечен в возрасте 50 суток. Ниже уровня деся тисуточного возраста количество эритроцитов в опытных группах не опускалось. Вместе с тем на протяжении всего исследуемого периода по ко личеству эритроцитов в крови цыплята опытных групп превосходят контрольных лишь в возрасте 40 суток на 7,64–26,56%. В остальных возраст ных группах исследуемый показатель у цыплят, получающих основной рацион, преобладает над опытными на 7,69–24,69%. Скорость оседания эритроцитов в возрасте десяти суток равна 2,3 мм/ч. У цыплят конт рольной группы максимума этот показатель достигает на 20е сутки и составляет 5,7 мм/ч. В первой опытной группе наибольшая скорость оседания эритроцитов отмечается в возрасте 50 суток (7,5 мм/ч). Во второй опытной группе максимальным этот показатель наблюдается у 20суточных цыплят и составляет 9,3 мм/ч. В течение всего опыта скорость оседания эрит роцитов в крови цыплят второй опытной группы больше, чем у цыплят контрольной, на 11,10– 111,10%. В первой опытной группе исследуемый показатель превышал аналогичный контрольной группы на 55,57–221,48%, за исключением 20суточного возраста, когда скорость оседания эритроцитов у цыплят, получающих «аминераль ный» рацион, на 23,54% меньше, чем у цыплят, получающих основной рацион. Количество лейкоцитов в возрасте десяти су ток составляет в среднем 5,0.109/л. В конт рольной группе количество этих клеток за ис следуемый период колеблется в пределах от 1,933.109/л до 5,80.109/л. Максимальный уро вень зафиксирован в возрасте 50 суток. В первой опытной группе количество лейкоцитов в различных возрастах составляет 1,667.109/л – 4,0 . 10 9/л, а во второй опытной группе – 1,40.109/л – 8,767.109/л. При этом наибольшими эти показатели отмечаются в возрасте 40 суток в группе с «аминеральным» кормлением и в 30 166 БИОЛОГИЧЕСКИЕ НАУКИ Лейкограмма цыплятбройлеров, % Группа цыплят Первая опытная Вторая опытная Контрольная Псевдоэозинофилы палочкоядерные сегментоядерные Эозинофилы Базофилы Лимфоциты Моноциты 1,0–5,0 1,3–3,3 2,0–4,0 40,7–52,7 40,7–62,0 40,7–58,3 1,0–3,5 0–4,0 0–2,0 0–1,0 0 0–1,0 36,7–52,0 30,7–46,7 34,3–52,0 3,3–7,3 2,3–7,3 3,3–10,7 суток у цыплят, получающих рацион с мини мальным содержанием минеральных веществ и обменной энергии. Доля различных форм лейкоцитов изменяется в зависимости от возраста и уровня кормления. Проанализировав возрастные изменения рас пределения доли различных форм лейкоцитов, лейкограмму соответствующей группы цыплят бройлеров можно представить следующим обра зом (табл.). Таким образом, уменьшение количества ми неральных веществ и обменной энергии в соста ве рациона цыплятбройлеров ведёт к снижению концентрации гемоглобина и количества эрит роцитов в крови, вместе с тем увеличивается скорость оседания эритроцитов. Перевод цып лят с основного рациона на новый уровень корм ления приводит сначала к снижению количества лейкоцитов на 13,76–27,57% по сравнению с контрольной группой, а в дальнейшем к увели чению этого показателя на 13,22–241,53% соот ветственно (p<0,001). Литература 1. Липунова, Е.А. Влияние новой полиминеральной кормо вой добавки на морфофункциональное состояние эрит роцитов периферической крови утят / Е.А. Липунова // Ус пехи современного естествознания. М., 2004. № 8. С. 120. 2. Фисинин, В.И. Рекомендации по кормлению с.х. птицы / В.И. Фисинин, И.А. Егоров, В.К. Менькин. М.: ВНИТИП МСХА, 2003. 143 с. Пути обогащения продукции птицеводства микроэлементами Т.В. Синюкова, к.б.н., ст. преподаватель, Оренбургский ГАУ Большинство регионов России, в том числе и Оренбургская область, являются йододефицит ными, в связи с чем зобная эндемия в нашей стране представляет собой весьма распростра ненное явление [1, 2, 3]. Одним из направлений в решении данной проблемы является увеличе ние на рынке таких продуктов питания, которые содержат повышенные количества данного мик роэлемента. Сегодня в пищевом рационе населе ния особое место занимают куриное мясо и яйца – натуральные диетические продукты, благопри ятно влияющие на здоровье. Обогащение пищевых яиц и мяса микроэле ментами, в том числе и йодом, – перспективное направление в науке и практике птицеводства. В основном такие пищевые продукты получают от курнесушек, включая в рацион различные кормовые добавки в дозах, превышающих реко мендуемые. Для получения йодированных яиц и мяса в качестве источника йода в комбикорм добавляют йодид калия. Однако получение такого продукта требует введения в рацион избыточного количества препарата йода, что может внести дисбаланс в функционирование нормальной микрофлоры пищеварительного тракта птиц и привести к ме таболической переориентации всего организма. При нарушении обмена веществ у птиц снижа ются продуктивность, резистентность, воспро изводительная функция. В условиях развития биотехнологий, в том числе и в области птице водства, возрастают экстремальные воздействия на птиц, поскольку по мере повышения продук тивности и ассортимента продукции они стано вятся более чувствительны к стрессовым факто рам, которые могут вызвать нарушения обмена веществ. Введение в рацион избыточного количества йодида калия является стрессовым фактором для организма курнесушек. Накопленный в лите ратуре научный материал позволяет считать, что микробный ответ в кишечнике при стрессовой ситуации является, в первую очередь, реакцией лактобактерий, включая нарушения экологичес кого барьера и колонизации условнопатоген ной микрофлоры [4, 5]. Кроме того, известно, что микрофлора желудочнокишечного тракта влияет на обмен йода, а именно: на процессы реабсорбции трийодтиронина (уменьшает поте ри йода с калом), осуществляя бактериальный гидролиз Т3конъюгатов (глюкоронидов и суль фатов) и участвуя в неферментативном пути по полнения пула трийодтиронина [6]. В связи с 167