Регуляция экспрессии генов у про- и эукариот
реклама

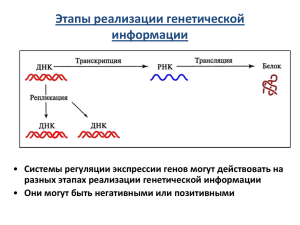
Тема: Регуляция экспрессии генов у про- и эукариот План лекции: 1. Общие принципы и механизмы регуляции активности генов. 2. Регуляция активности генов у прокариот. 3. Регуляция активности генов у эукариот. 4. Уровни регуляции активности генов и их характеристики Регуляция активности (экспрессии) генов позволяет приспособить организм к изменяющимся условиям среды. В клетке в каждый момент ее существования должно синтезироваться и функционировать необходимое количество различных белков. У бактерий набор белков регулируется наличием м-РНК, пригодных для биосинтеза белка. Так как процессы транскрипции и трансляции протекают почти одновременно, то внести какиелибо изменения в структуру м-РНК практически невозможно. У эукариот молекулы РНК подвергаются существенным модификациям благодаря ядерной мембране, разделяющей зоны транскрипции и трансляции. В любом случае первым и главным элементом контроля реализации генетической информации от ДНК к белку у всех организмов является контроль на уровне транскрипции. Регуляция экспрессии у прокариот Регуляция генетической активности у прокариот идет на уровне транскрипции. В 1961 г. франц. ученые Жакоб и Моно предложили модель оперона как систему регуляции экспрессии генов бактерий. Оперон Промотор- место связывания РНК-П Оператор- место связывания белкарепрессора Структурные гены- кодируют ферменты -------------------------------------------------------Ген-регулятор кодирует белокрепрессор. Он не входит в состав оперона. Индуцибельные опероны Регулятором является исходный субстрат (S) В отсутствие этого субстрата белокрепрессор имеет высокое сродство к оператору, поэтому РНК-П не может начать транскрипцию (оперон выключен). При накоплении метаболита (S) в клетке некоторое его количество связывается с белком-репрессором и понижает его сродство к оператору. Оперон включается и синтезируются ферменты для расщепления S. Репрессибельные опероны Регулятором служит конечный продукт (Р) В отсутствие этого продукта белокрепрессор имеет низкое сродство к оператору, поэтому РНК-П транскрибирует гены оперона. Оперон включен и синтезируются ферменты, способствующие образованию вещества Р. При накоплении данного в-ва некоторое его кол-во связывается с белком-репрессором и повышает его сродство к оператору. Оперон выключается и синтез ферментов прекращается. Способы регуляции экспрессии генов прокариот Регуляция связывания РНК-П с промоторами Регуляция перемещения связавшейся РНК-П от промотора к собственно генам Лактозный оперон Лактоза- дисахарид, на котором растут клетки бактерий Lac Z – галактозидазу для расщепления лактозы до глюкозы и галактозы Lac Y- пермеазу для проникновения лактозы из внешней среды в клетку Lac A - трансацетилазу Организация и функционирование Lac – оперона по Жакобу и Моно Организация и функционирование Lac – оперона по Жакобу и Моно Функционирование лактозного оперона у E. Coli: а– локализация сайтов связывания молекул РНК – полимеразы и репрессора в регуляторный области гена lac Z [ Lewin, 1994. P. 417]; б – структура лактозного оперона (как и у всех генов, регулируемых САР и сАМР, промотор содержит два района: участок связывания с РНК-полимеразой и участок связывания с комплексом САР и сАМР); в – негативная, г – позитивная регуляция lac-оперона [ б-г – Curtis,Barnes, 1989. P. 325]. В лактозном опероне левая часть промотора (активатор) связывается с белком-активатором катаболизма (БАК, или САР в англ.терминологии), а правая часть – с РНК-П. CАP-белок в отличие от белкарепрессора играет позитивную роль, помогая РНК-П начать транскрипцию. Связывание CАP с промотором происходит если CАP соединяется с цАМФ. Глюкозы нет Если источником энергии служит лактоза и во внешней среде нет глюкозы, репрессор инактивируется. При этом промотор открыт для РНК-П, транскрипция идет. . Глюкоза есть Если во внешней среде достаточно глюкозы, то биологического смысла в использовании лактозы уже нет. Поэтому глюкоза препятствует активации лактозного оперона даже в присутствии больших количеств лактозы. В отсутствие глюкозы В отсутствие глюкозы в клетке уровень цАМФ возрастает (сигнал голода) , поэтому CАP связан с лактозным промотором, к которому легко присоединяется РНК-П. В этих условиях активность оперона зависит только от того, свободен или нет оператор,т.е. от наличия во внешней среде лактозы. В присутствии глюкозы Уровень цАМФ остается низким, промотор остается без CАP и теряет сродство к РНК-П. Лактозный оперон не работает и в качестве источника питания используется глюкоза. Триптофановый оперон Это репрессибельный оперон с двойным механизмом регуляции: 1.регулируется перемещение РНК-П по оператору 2.окончание транскрипции на аттенюаторе Т.О. состоит из 5 цистронов. За промотором и оператором находится лидерный отдел, который заканчивается аттенюатором. При транскрипции этого отдела образуется лидерный участок мРНК. При трансляции образуется лидерный пептид. Особенность лидерного пептида: среди его 14 аминокислотных остатков содержатся 2 остатка триптофана, т.е. той самой аминокислоты, синтез которой контролируется опероном. Триптофан есть Синтез лидерного пептида идет без задержки. При достижении РНК-П аттенюатора срабатывает сигнал об окончании транскрипции. Триптофана нет Синтез лидерного пептида задерживается и сигнал об окончании трансляции на аттенюаторе не срабатывает и синтезируются ферменты, необходимые для пополнения запаса триптофана в клетке. Аттенюатор Это участок ДНК между оператором и генами, на котором при определенных условиях прекращается транскрипция оперона. За промотором и оператором находится лидерный отдел, именно он оканчивается аттенюатором. Когда триптофана мало Синтез ЛП задерживается и сигнал об окончании транскрипции на аттенюаторе не срабатывает, поэтому синтезируются ферменты, необходимые для пополнения запаса триптофана в клетке. Координированная репрессия у Salmonella typhimurium Литература: 1. Биология. Под ред. Ярыгина В.Н. М., 2001. 2. Бочков Н.П. и др. Медицинская генетика. М., 1984. 3. Генетика. Под ред. Иванова В.И. М., 2006. 4. Заяц Р.Г. и др. Общая и медицинская генетика. Ростов-на-Дону, 2002. 5. Инге-Вечтомов С.Г. Генетика с основами селекции. М., 1989. 6. Кемп П., Армс К. Введение в биологию. М., 1988. 7. Медицинская биология и генетика. Под ред. проф. Куандыкова Е.У. Алматы, 2004. 8. Фаллер Д. М., Шилдс Д. Молекулярная биология клетки. М., 2006. 9. Муминов Т.А., Куандыков Е.У. Основы молекулярной биологии (курс лекций). Алматы, 2007. 10. Мушкамбаров Н.Н., Кузнецов С.Л. Молекулярная биология. М., 2003. Контрольные вопросы (обратная связь): 1. Что такое оперон? 2. Уровни регуляции активности генов у прокариот. 3. Механизмы регуляции активности генов у прокариот. 4. Особенности регуляции активности генов у эукариот. 5. Дифференциальная экспрессия генов и ее значение в жизнедеятельности организмов.