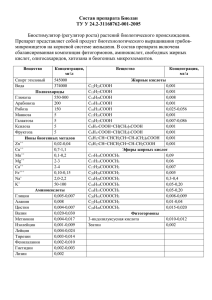
Роль желчных кислот в переваривании жиров. Хиломикроны
реклама

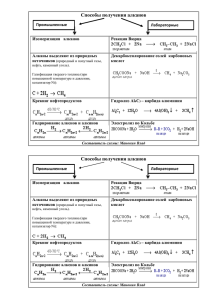
Катаболизм жирных кислот. Образование кетоновых тел. Катаболизм триацилглицеролов, -окисление жирных кислот В период изобилия пищи излишек углеводов превращается в жир. Когда настают менее удачные дни, организм мобилизует свои жировые запасы. Жир из депо, обычно подкожной области, транспортируется в печень и другие органы, где он окисляется, выделяя при этом требуемую для нормального существования энергию. Первая стадия распада жиров – гидролиз триацилглицеролов, содержащихся в хиломикронах и липопротеинах низкой плотности, катализируемый ферментом липопротеинлипазой. В результате образуются глицерол и свободные жирные кислоты. • Глицерол далее превращается очень просто. Подвергается фосфорилированию по одному боковому гидроксилу. Образовавшийся глицерол-3-фосфат окисляется до дигидроксиацетонфосфата. Он включается в реакции расщепления углеводов до СО2 и Н2О, либо глюконеогенеза, обеспечивая синтез глюкозы. Окисление жирных кислот протекает после подготовки, на которую необходимо затратить энергию. Энергия поставляется за счет АТФ, но при подготовке одной молекулы жирной кислоты расходуется сразу две макроэргические фосфатные связи АТФ, которая превращается в АМФ. • Энергия от АТФ к жирной кислоте переходит не за счет фосфорилирования молекулы субстрата, а путем образования продукта взаимодействия жирной кислоты с коэнзимом А. Этот процесс катализирует фермент ацил-КоА-синтетаза. • Жирные кислоты с длинной цепью от 12 до 20 атомов углерода активируются на внешней мембране митохондрий, а с короткой и средней длиной от 4 до 12 атомов углерода – в матриксе митохондрий, куда они проникают путем диффузии. Транспорт ацил-КоА жирных кислот с длинной цепью происходит путем взаимодействия с карнитином и с помощью двух ферментов (карнитин-ацил-трансфераз). β-окисление жирных кислот 1. Активация жирных кислот R-CH2–CH2 –COOH + HS~KoA + АТФ R-CH2-CH2-CO~S-KoA + АМФ + ФФ ацил~КоА Ацил-КоА-синтетаза (тиокиназа) МАТРИКС наружная мембрана внутренняя мембрана H3C CH3 HO CH CH2 R-C~S-KoA C O O карнитинацилтрансфераза I H3C HS-KoA H3C N CH3 Т р а н с л о к а з а HO CH CH2 CH2 O H3C H3C N CH3 H2C O CH CH2 O C O R-C~S-KoA карнитинацилтрансфераза II C O O O R-C O CH O C H2C R-C CH3 H2C H2C O N H3C N H3C H3C O O HS-KoA β-окисление жирных кислот Вначале происходит окисление под действием дегидрогеназы флавопротеинового ряда с образованием двойной связи между - и -атомами углерода. β O H3C-(CH2)n -CH2-CH2-CH2-C~SKoA ФАД ацил-КоА-дегидрогеназа 1,5 АТФ ФАД-Н2 β O H3C-(CH2)n -CH2-CH=CH-C~SKoA Еноил-КоА Ацил-КоА β-окисление жирных кислот O β H3C-(CH2)n -CH2-CH=CH-C~SKoA Еноил-КоА Н2О еноил-КоА-гидратаза ОН O H3C-(CH2)n -CH2-CH-CH2-C~SKoA β β-гидроксиацил-КоА β-окисление жирных кислот ОН O H3C-(CH2)n -CH2-CH-CH2-C~SKoA β-гидроксиацил-КоА β НАД НАДН+Н+ βО β-гидроксиацил-КоАдегидрогеназа 2,5 АТФ O H3C-(CH2)n -CH2-C -CH2-C~SKoA β-кетоацил-КоА β-окисление жирных кислот βО O β-кетоацил-КоА H3C-(CH2)n -CH2-C -CH2-C~SKoA HS-KoA β-кетоацил-КоА-тиолаза O H3C-(CH2)n-2 -CH2-C~SKoA Ацил-КоА с=(n-2) Новый ацил-КоА на 2 атома С короче исходного ацила, который окисляется далее по той же схеме -окисления. O CH3-C~S-KoA ЦТК Ацетил-КоА c=2 Если ацетил-КоА поступает в цикл Кребса, то распадается до СО2 и Н2О, обеспечивая синтез ~10 M АТФ на 1 М ацетил-КоА. β-окисление жирных кислот •β-Окисление пальмитоил-КоА •Количество молекул АТФ •7 NADH (от пальмитоил-КоА до ацетилКоА), •окисление каждой молекулы •кофермента в ЦПЭ обеспечивает •синтез ~2,5 АТФ • 17,5 •7 FADH2, окисление каждой молекулы •кофермента в ЦПЭ обеспечивает синтез • ~1,5 АТФ •10,5 •8 молекул ацетил-КоА •в ЦТК каждая молекула обеспечивает синтез ~10, АТФ Суммарное количество молекул АТФ, синтезированных •при окислении одной молекулы пальмитоил-КоА •80 •108 β-окисление жирных кислот с нечетным числом атомов С С9 O CH3-C~S-KoA С7 O CH3-C~S-KoA С5 O CH3-C~S-KoA С3 O H3C H2 C C S-KoA пропионил-КоА +СО2 О HOOC-(CH2)2-C~SKoA ЦТК сукцинил-КоА β-окисление ненасыщенных жирных кислот О H3C-(CH2)7 -CH=CH-CH2-C~S-KoA ∆3,4 –еноил-КоА ∆3,4 – цис - ∆2,3 – трансеноил-КоА-изомераза О H3C-(CH2)7 -CH2-CH=CH-C~S-KoA ∆2,3 –еноил-КоА Н2О еноил-КоА-гидратаза аналогично β-окислению насыщенных жирных кислот со стадии гидратации Синтез кетоновых тел в митохондриях печени из ацетил-КоА. O Кетоновые тела 2 H3C C SKoA HS-KoA HS-KoA O H3C C H2O тиолаза O H2 C C SKoA ацетоацетил-КоА O ГМГ-Ко-синтаза H3C C SKoA HS-KoA O H2 O C C OH C H2 C O C SKoA CH3 O H3C C SKoA β-гидрокси-β-метилглутарил-КоА ГМГ-КоА-лиаза Кетоновые тела O H2 O C C OH H2 C C O C SKoA β-гидрокси-β-метилглутарил-КоА CH3 O H3C C SKoA O H3C CO2 C H2 C ГМГ-КоА-лиаза O ацетоацетат C O НАД Н НАД H+ O H3C C ацетон CH3 H3C H C H2 C OH β-оксибутират O C O H3C H C O H2 C C O OH β-оксибутират НАД Н НАД НАД Н H+ O H3C О сукцинил-КоА H3C C O C O O H2 C ацетоацетат Сукцинил-КоА-ацетоацетаттрансфераза HOOC-(CH2)2-C~SKoA O C H2 C C SКoA ацетоацетил-КоА O H3C C O H2 C C SCoA тиолаза +HS-KoA O 2 H3C C SKoA ацетоацетил-КоА • Образование каждой молекулы ацетоацетата, сначала накапливающегося в печени, а затем выбрасываемого из нее в кровь, означает, что две молекулы ацетил-КоА с запасенной в них энергией не входят в цикл Кребса. • Причина этого лежит в схеме работы этого цикла. Ацетил-КоА должен реагировать с оксалоацетатом и давать цитрат. Оксалоацетат образуется из пирувата. Пируват продукт метаболизма глюкозы, а не жирных кислот. Окисление же жирных кислот в значительном количестве осуществляется как раз в условиях углеводного голодания. По мере того как в митохондриях печеночных клеток возрастает концентрация высокореакционноспособного ацетилКоА, все большее число их попарно реагирует друг с другом. • Когда отсутствуют углеводы, клетка вынуждена искать источник энергии в жировых запасах. Но она оказывается не в состоянии использовать это горючее столь эффективно, как глюкозу. • Этот пример позволяет еще раз подчеркнуть важность углеводов для энергетического обмена веществ. • Некоторое количество кетоновых тел образуется постоянно. При длительном голодании кетоновые тела становятся основным источником энергии для скелетных мышц, сердца и почек. Через 2-3 дня голодания они проникают в клетки мозга, окисляются там, чем снижают их потребность в глюкозе.