Tsitologia_3x
реклама

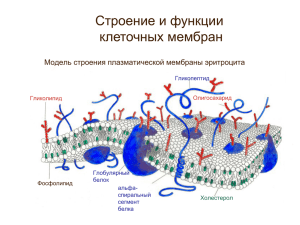
Животные и растительные клетки эукариот практически не отличаются. Они состоят из органических веществ, неорганических и только очень тонкие иммунологические что-то там позволят нам отличит клетки животных от растений. Так же существуют различия. Растительная клетка способна связывать углекислоту в процессе фотосинтеза. Растительная клетка имеет жесткую клеточную стенку, в состав которой входит целлюлоза. Растительная клетка имеет два типа органоидов, которых нет у животных клеток. Это пластиды трех типов и центральная вакуоль. Метаболизм в растительных и животных клетках прямо противоположен. Резервным углеводом растительных клеток является полисахарид крахмал, а в животных клетках и клетках грибов запасной углевод – гликоген. Практически у всех растительных клеток и у клеток грибов отсутствуют структуры немембранного происхождения, состоящие из микротрубочек. Это центриоли и базальные тельца. Также будут принципиальные отличия практически по каждому органоиду. Цитоплазма и строение биомембран Цитоплазма эукариотических клеток имеет следующий состав: вода занимает 80%, белок около 10%, ДНК 0,4%, РНК 0,7%, липиды около 2%, неорганические соли 1% и остальные соли 1%. Цитоплазма – внутренний компонент клетки без ядра. Строение цитоплазмы подразделяют на три части: органоиды, включения и гиалоплазма (основная жидкая фаза клетки – цитозоль). Органоиды – обязательные для любой клетки компоненты. Включения – необязательные компоненты, которые являются либо отложениями запасных веществ, либо скоплением продуктов метаболизма. Гиалоплазма (лат. – прозрачный) – истинная внутренняя среда клетки. Сложный состав. По консистенции приближается к гелю. Гели – структурированные коллоидные системы с жидкой дисперсной средой. Частицы дисперсной фазы соединены между собой в рыхлую пространственную структуру, что лишает систему текучести. Гель цитоплазмы относится к тиксотропным гелям, которые под воздействием внешних условий, которые могут менять свое агрегатное состояние и переходить в менее вязкую фазу, которая называется золь. Организуют цитоплазму структуры цитоскелета. Наибольшая роль принадлежит микротрубочкам. Гиалоплазма выполняет: 1) Синтез и отложение запасных полисахаридов, накопление липидов. 2) Место протекания гликолиза и синтеза АТФ. 3) Место биосинтеза. Активация аминокислот с помощью специфических ферментов и связывания их с транспортными РНК. 4) В гиалоплазме происходит модификация ферментов, которая приводит к изменению функций и структуры белков. 5) Место локализации всех строительных блоков биомембран, а также всех промежуточных метаболитов. 6) Локализация всех неорганических соединений, причем концентрация неорганических веществ строго детерминирована и регулируется органоидами клетки. Все без исключения клеточные мембраны построены по общему принципу: тонкие липопротеидные пленки, состоящие из двойного слоя липидных молекул, в который включены молекулы белка. В весовом отношении, в зависимости от типов мембран, на долю липидов приходится от 25% до 60%, а на долю белков от 40% до 70%. Углеводы – необязательный компонент биомембран. От 2% до 10%. Внутренние мембраны в клетке замыкаются сами на себя, таким образом, что образуют полости и тем самым разделяют цитоплазму на отсеки. Внутреннее их содержание всегда отличается от содержимого гиалоплазмы. Толщина – 7 – 10нм. По массе около 4%. По общей площади внутренние мембраны существенно превышают наружную мембрану – плазмолемма. Структурной основой биологических мембран всегда является двойной слой липидов. К липидам относится большая группа веществ, обладающих плохой растворимостью в воде и хорошей растворимостью в органических соединений. Характерными липидами, формирующими бислой, являются фосфолипиды. Также существуют стероидные липиды и сфингомиелины. Фосфолипиды = глицерофосфопиды = глицеролипиды. Сложные эфиры глицерина с тремя жирными кислотами и с фосфорной кислотой. Сфингомеилины отличаются тем, что глицерин замещен сфингоспиртом. Характерной особенностью липидов является разделение их молекул на две функционально различные части – неполярную часть (хвостовая часть) и полярная (головная часть). Полярные головки имеют либо отрицательный, либо нейтральный. Наличие хвостиков объясняет хорошую растворимость липидов в жирах и неорганике. Если полярные липиды смешать с водой, то образуется эмульсия. При этом гидрофобные участки будут стараться образовать однородную фазу. Если к липидам добавить немного воды, гидрофобные хвосты, мицеллы вывернуты наизнанку и гидрофобные хвосты будут торчать наружу. Самопроизвольное образование бислоя. Качественный и количественный состав липидов мембран различен: плазматические мембраны клеток животных богаты холестерином. Примерно до 30%. Очень мало лецитина. Но внутренние мембраны богаты фосфолипидами и бедны холестерином. В целом, для плазматической мембраны характерно преобладание насыщенных и мононенасыщенных жирных кислот в составе фосфолипидов, тогда как во внутренних мембранах наоборот – много полиненасыщенных жирных кислот. Поэтому внутренние мембраны менее жестки, чем плазматические. В 1925 году Грендель и Гортер показали, что липиды – основа мембран. Мембраны х непроницаемый барьер для любых заряженных молекул. Свободные диффузии через бислой мембран встречаются редко. 1935 год. Даниэль и Даусен внесли дополнение к модели липидов, отметив, что в состав мембран входят белки. Бутербродная модель. По их представлению, белки располагались сплошными рядами в мембранах. 1980 год. Бутербродная мембрана встречается редко. 1971 год. Сингер и Николсон. Современная модель. Жидкостно-мозаичная модель. Согласно ей, мембрана состоит из неплотно упокованых белковых глобулярных структур, свободное пространство между которыми заполнено липидными молекулами. Большая часть липидных молекул не связана с белками. Белковые молекулы как бы плавают в липидном слое. Так как липидные молекулы маленькие и легкие, то их число по отношению к белкам всегда выше раз в 50. Связь между липидами и белками осуществляется с помощью солевых и ионных связей. Большая часть белком взаимодействует с липидами на основе гидрофобных взаимодействий. Мембранные белки, которые встраиваются в бислой также в своей молекуле имеют гидрофобные и гидрофильные участки. Такой белок относится к интегральным белкам и очень жестко взаимодействует с липидами. Размер интегральных белков равен 8 нм. Встречаются до 35нм. Обычно интегральные белки ассиметричны по своей природе. С цитоплазматической стороны интегральные белки связываются с периферическими белками. Они обеспечивает взаимодействие мембран с субмембранными структурами. Полуинтегральные – погружены лишь на половину. Белки разделяют на три группы: 1) Ферменты; 2) Рецепторы; 3) Структурные белки. Набор ферментов в составе мембраны довольно разнообразен. В плазмолемме и где-то там обнаружено до 34 молекулах. В митохондриях специфическим набором ферментов является транспортная цепь электронов. АТФ-синтетаза. Рецепторные белки – специфически связываются с веществами, узнают их и обеспечивают в случае клеточной потребности проведение их внутрь клетки. И белки и липиды обладают подвижностью. Липиды двигаются постоянной с огромной скоростью – 2 мкм\с. Они могут двигаться вдоль липидного слоя.Латеральная диффузия. Могут вращаться вокруг своей оси.вращательная диффузия. Могут переходить из внутреннего слоя во внешний. Флип-флоп. Белки могут перемещаться латерально, но скорость их ниже. Плазмолемма выполняет следующие функции: 1) Барьерная. Плазмолемма служит преградой между сложноорганизованным внутриклеточным содержимым и внешней средой. Механическую устойчивость плазмолеммы дополнительно обеспечивают примыкающие к ней со стороны цитоплазмы в структуре цитоскелета и кортикального слоя цитоплазмы. В кортикальном слое (0,1 – 0,5мкм) отсутствуют рибосомы и мембранные пузырьки. Сосредоточено большое количество микрофиламентов и микротрубочек. Причем, в компонентах микрофибрилярной системы преобладают актиновые фибриллы. Они участвуют в сократительных движениях плазмолеммы и заякоривании белков. Плазматическая мембрана участвует в формировании клеточных стенок. Участвует в образовании пелликулы, жесткого слоя простейших и инфузорний. Барьерная роль плазмолеммы заключается в ограничении свободной диффузии веществ. Мембраны проницаемы для воды, газов, малых неполярных молекул жирорастворимых веществ, но совершенно не проницаемы для заряженных ионов и для крупных биополимеров. рецепторная функция связана с узнаваемостью различных химических веществ. В качестве рецептора узнающих структур выступают элементы надмембранной структур в животных клетках - гликокаликс. Рецептурными компонентами являются гликопротеины. Гликокаликс в животных клетках имеет толщину 3 – 4 нм.и обеспечивает ряд ферментативных реакций, например, в мембранах тонкого кишечника внеклеточное пищеварение. Клетки животных могут обладать разным набором рецепторов или разной чувствительностью одного и того же рецептора. Роль рецептора выражается связанностью со специфическими веществами и способностью реагировать на физические раздражители. Кроме того, рецепторы передают межклеточные сигналы с поверхностью клетки внутрь цитозоли. Самой изученной является система передачи сигнала клетки с помощью гормонов. За счет аденилат-циклазной системы, которая служит преобразователем внешнего сигнала. 1) Гормон взаимодействует специфически с рецептором мембраны. Не проникая внутрь клетки активирует фермент аденилат-циклазу, которая синтезирует циклическую АМФ в свою очередь, активирующий внутриклеточный фермент. Таким образом, сигнал от плазмолеммы передается внутрь клетки. Разнообразие рецепторов на плазмолемме приводит к созданию сложной системы, которая позволяет отличать одни клетки от других. Сходные клетки вступают во взаимодействие, приводящее к слипанию к поверхности. С плазмотической мембраной связана локализация специфических рецепторов, реагирующих на физические факторы -= например, в плазмолемме растений локализованы белкирецепторы, взаимодействующие с квантами.света. В плазмолемме мембран светочувствительных животных расположены система фоторецепторынх белков. Эти ферменты обеспечивают преобразование световой энергии в химическую энергию и т.д. Транспортная функция. Осуществляет регулируемый избирательный транспорт веществ и выполняет роль первичного клеточного анализатора. Плазмолемма является полупроницаемой. Через нее с различной скоростью проходят разные молекулы. Максимальной проникающей способностью обладает вода и растворенные в ней газы. Ионы проникают медленно. Скорость воды 10^-4 см в секунду. В мембране существуют для этого специальные поры. Число их составляет примерно 0,06% всей клеточной поверхности. Плазматическая мембрана способна транспортировать ионы и многие мономеры, такие как сахара, аминокислоты. Транспорт ионов через мембрану происходит за счет специальных транспортных белков, которые называются пермеазы. Эти белки могут везти транспорт либо в одном направлении одного вещества. Унипорт Либо в нескольких веществ одновременно. Симпорт. Иил транспорт двух веществ в противоположенных направлениях. Антипорт Пасс. Транспорт. Так, например, в клетку проникают ионы натрия из внешней среды, где концентрация их выше, чем в цитоплазме.В случае пассивного транспорта некоторые мембранные транспортные белки образуют молекулярные комплексы (каналы) через которые растворенные молекулы проходят через мембрану за счет простой диффузии. Часть этих каналов открыта потоянрно, другая часть этих каналов может закрываться и открываться либо на связывание с сигнальными молекулами либо на изменение внутриклеточной концентрации. Специальные транспортные белки избирательно связываются с тем или иным ионом и проходят через его мембрану. Это облегченная диффузия. В клетках существуют и мембранные белковые переносчики, которые работают против градиента концентрации. Затрачивается энергия АТФ. Такой тип транспорт называется активным транспортом. Он осуществляется с помощью белковых ионных насосов. В плазмотической мембране находится двухсубъединичная молекула, которая одновременно является и чем-то еще. К-АТФ-аза. Откачивает за один цикл три иона натрия из клетки, а внутрь закачивает два иона калия против градиента концентрации. Затрачивается одна АТФ, которая идет на реакция фосфолирирования. В результате натрий переносится из клетки, а калий получает возможность транспортироваться внутрь клетки. В результате активного транспорта гомеостаз характеризуется удивительным постоянством. Также могут переноситься ионы магния и кальция. 80% АТФ расходуется на поддержание гомеостаза. Также транспорт осуществляется с помощью везикулярного переноса. Два типа транспорта: эндоцитоз и экзацитоз. При эндоцитозе определенные участки плазмолеммы захватывает и как бы обволакивает внеклеточный материал, заключая его в мембранную вакуоль, возникшую за счет впячивания плазматической мембраны. Белок клатрин - маркер везикулы. Эндосома. Она может иметь два типа: фагосома и пиносома. Эндоцитоз формально разделяют на пиноцитоз и фагоцитоз. Эндоцитоз – основной механизм получения строительных блоков за счет внутриклеточного переваривания, которое осуществляется на втором этапе эндоцитоза после слияния первичной эндосомы с лизосомой, вакуолью, содержащей набор гидролитических элементов или гидролаз. Эндоцитоз был писан Мечниковым. Эндоцитоз включает пиноцитоз и фагоцитоз. Может быть неспецифическим или конститутивным (потоянным), постоянным и специфическим (рецепторным). Неспецифический эндоцитоз протекает автоматически и часто приводит к захвату и поглощению ненужных клетке веществ. Сопровождается первоначальной сорбцией гликокаликса. Гликокаликс имеет кислые группировки на углеводной части. Имеют отрицательный заряд. За счет этого предпочитительное электростатическое взаимодействие. При таком адсорбционном неспецифическом эндоцитозе поглощаются макромолекулы и мелкие частицы (кислые белки, антитела, коллоидные частицы). Жидкофазный пиноцитоз приводит к поглощению растворимых в жидкой среде молекул, которые не взаимодействуют с плазмолеммой. На следующем этапе происходит изменение морфологии клеточной поверхности. В начале образуются окаймленные ямки, состоящии из основного белка какого-то. Три его молекулы вместе с тремя молекулами низкомолекулярного вспомогательного белка образуют рыхлую сеть, состоящую из пяти и шестиугольных структур. Клатриновый слой окаймляет весь периметр отделяющихся первичных эндоцитозных вакуолей и обеспечивает их слияние и последующий транспорт в клетке. При поступление внутрь цитозоля клатриновый слой распадается, диссоциирует и мембрана эндосомы приобретает обычный вид. После потери клатринового слоя эндосомы сливаются друг с другом. Мембраны окаймленных ямок содержат сравнительно мало холестерина и за счет процессов сливания и объединения слоев контактирующих мембран образуется эндосома. В ходе фагоцитоза и пиноцитоза клетки теряют большую площадь плазмолеммы, которая довольно быстро востанавливается за счет процесса рециклизации мембран. Это процесс возвращения вакуолей и их встраивания в плазмолемму. Таким образом при рециклизации мы имеем дело с челночным механизмом переноса мембран. Плазмолемма – эндосома – вакуоль – плазмолемма. Специфический или опосредуемый рецепторами эндоцитоз отличается тем, что на мембране имеются специфические рецепторы – лиганды.