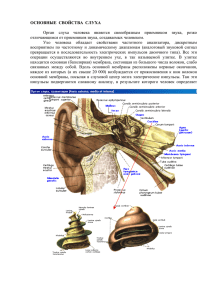
С. Б. Парин ОСОБЕННОСТИ БИОЭЛЕКТРОГЕНЕЗА. ИНФОРМОНЫ. МАЛЫЕ НЕЙРОННЫЕ СЕТИ Принципы переработки информации в нервной системе
реклама

Российская академия наук Институт прикладной физики Научно-образовательный центр ИПФ РАН С. Б. Парин ОСОБЕННОСТИ БИОЭЛЕКТРОГЕНЕЗА. ИНФОРМОНЫ. МАЛЫЕ НЕЙРОННЫЕ СЕТИ Принципы переработки информации в нервной системе Уч еб но -м ето дич еско е по со б ие по ку рсу « Нейро фи зио л о гия» Нижний Новгород Издательство ИПФ РАН 2022 УДК 573 ББК 28.707.3 Рецензенты К.В.Анохин, академик РАН, доктор медицинских наук, зав. лабораторией нейробиологии памяти Научно-исследовательского института нормальной физиологии им. П.К.Анохина РАН; Д. Б. Гелашвили, доктор биологических наук, профессор, зав. кафедрой экологии биологического факультета ННГУ им. Н.И.Лобачевского; В. Г. Яхно, доктор физико-математических наук, зав. лабораторией автоволновых процессов ИПФ РАН. П18 Парин Сергей Борисович Особенности биоэлектрогенеза. Принципы переработки информации в нервной системе: Учебнометодическое пособие. ― Нижний Новгород: Изд-во ИПФ РАН, 2022. ― 72 с. ISBN 978-5-8048-0068-1 Данное учебно-методическое пособие обобщает сорокалетний опыт чтения автором различных нейробиологических курсов студентам технических, биологических, медицинских и психологических специальностей. В максимально образной форме описаны базовые принципы передачи информации в нервной системе с помощью электрических сигналов. Предназначено для представителей физико-математических специальностей, а также психологов, биологов и медиков, изучающих введение в механизмы переработки информации в нервной системе и желающих разобраться в основах биоэлектрогенеза в нервных тканях, то есть начинающих свой путь в современной нейробиологии. ISBN 978-5-8048-0068-1 © Институт прикладной физики РАН, 2022. © Парин С. Б., 2022. ВВЕДЕНИЕ Книжка, которую Вы держите в руках, написана как своеобразный самоучитель для тех, кто хочет разобраться в основах биоэлектрогенеза в животных тканях (прежде всего, в нервной системе животных). Она не претендует на глубину анализа, у неё совсем другая задача: дать начинающим свой путь в нейробиологии физикам, химикам, психологам, медикам и (как это ни грустно) биологам стартовую площадку, базу для дальнейшего погружения. Думаю, Вы уже успели заметить, что учебная литература бывает двух типов. По первому типу написаны немногочисленные (в основном, увы, неотечественные) учебники, авторы которых просто и внятно описывают те сложные закономерности, в выявлении которых им посчастливилось участвовать самим. В таких книжках легко дышится и продуктивно думается. Второй тип представляют бесконечно переписываемые из издания в издание нагромождения фактов и гипотез, многие из которых по ходу дела возводятся в ранг фундаментальных теорий, каковыми ни в коей мере не являются. Эти книжки опасны для доверчивого читателя, потому что создают иллюзию всепознанности и становятся питательной средой для всяческого парарелигиозного мракобесия. «Учёные показали…» ― эта очень страшная фраза переселилась в многочисленные рекламные клипы именно из вполне наукообразной учебной литературы. Нейробиологии в этом смысле, казалось бы, повезло. Есть прекрасные отечественные издания, написанные признанными в мире классиками «жанра»: это и многократно переиздававшиеся учебники П.Г. Костюка, и книжки П.К. Анохина, А.Б. Когана, В.В. Шульговского, Б.И. Ходорова, А.Д. Ноздрачёва, О.А. Крышталя и многих других, и известные нам благодаря переводам труды Э. Кэндела, К. Прибрама, Ф. Блума, В. Маунткастла, Д. Эдельмана, Г. Шеперда, Д. Экклза, С. Куффлера… Казалось бы, бери и читай! Замечательно, но есть одно «НО»: эти прекрасные книги рассчитаны на уже подготовленного читателя. Значит, есть такая задача – подготовить заинтересованного читателя (независимо от его первоначального образования) к их восприятию. Автор ни в коем случае не гарантирует абсолютной доказанности всех изложенных в пособии научных фактов (здесь найдётся место и гипотезам), однако обещает приложить все усилия, чтобы отделить то, что реально промерено, от того, что является результатом игр разума, пусть и очень красивых. И ещё: я постараюсь не пугать читателя страшными словами там, где можно обойтись простым человеческим языком. Мне хотелось, чтобы получился спокойный разговор с читателем. В конце концов мы ведь будем говорить о том, что смогли открыть, увидеть люди, пытаясь заглянуть в наш мозг (рис. 1). Неужели вместе мы не сможем это понять? Рис. 1. Так выглядел бы наш мозг, если бы мы могли видеть его насквозь. Впрочем, современные томографические методы позволяют это проделывать (пусть и путём компьютерной реконструкции). Завершая введение, хочу поблагодарить заведующую кафедрой психофизиологии ННГУ, доктора биологических наук Софью Александровну Полевую за помощь на всех этапах выполнения этой работы. ПОНЯТИЕ БИОЭЛЕКТРОГЕНЕЗА. ВОЗБУДИМОСТЬ. ВОЗБУДИМЫЕ КЛЕТКИ Мир, в котором мы все живём, бескрайне многолик и изменчив. Его лица и изменения могут быть прямо обращены к нам, но могут существовать и независимо от нас. В любом случае, как только мы хоть краешком своего Я прикасаемся к этому миру, мы входим с ним во взаимодействие, и должны иметь устройства (аппараты, приборы, системы…), обеспечивающие эффективность этих взаимодействий. первым и главным из этих устройств является наша нервная система. Её задача — собирать информацию из внешнего мира, обрабатывать её и, сопоставив её с накопленным раньше опытом, принимать управленческие решения: как реагировать, и реагировать ли вообще. То есть наша нервная система — это информационная система со всеми её атрибутами. А раз так, должен быть язык, код, правила, по которым записывается информация. Вот мы и добрались до биоэлектрогенеза, до особенностей возникновения электрических процессов в живых системах. Подозрение о том, что базовым, первичным кодом в нервной системе (подобно двоичному коду любого компьютера, где 00, 01, 11 или 10 удивительным образом ухитряется вместить все помыслы писателя, поверяемые им машине) может быть импульсный код, то есть определённая последовательность нервных (по сути, электрических) импульсов, возникло не на пустом месте. Ещё в конце XVII века в ожесточённом споре двух великих физиков — Луиджи Гальвани и Александро Вольта — родилась первая истина на этом долгом пути: в теле животных (и человека, естественно) есть электрические потенциалы и электрический ток. Потребовалось два с лишним века, чтобы мы начали понимать, что именно эти биоэлектрические явления и дают возможность нервной системе работать с информацией, и ещё почти век, чтобы начать понимать, как это происходит. Давайте обсудим, что мы сегодня знаем об этом. Есть устойчивое мнение, что способностью к биоэлектрогенезу обладают только некоторые клетки животных, которые принято называть возбудимыми. Честно говоря, это не совсем правда! Сегодня уже хорошо известно, что электрический заряд на мембране имеют и клетки крови (например, эритроциты), и гепатоциты (то есть клетки печени), и половые клетки животных, и т.д. Более того, некоторые клетки растений тоже способны к генерации электрического заряда на своей поверхности, причём этот заряд меняется и распространяется, что позволяет биофизикам уже полвека говорить о наличии у растений нервной системы (тех, кто, в отличие от ботаников, наблюдал растения в реальной жизни, а не в гербарии, это едва ли удивит). Тем не менее, клетки нервной, эндокринной и мышечной систем традиционно относят к особой группе, называя их (и только их) возбудимыми. Почему же? Ответ начнём с формулирования очень важного в нейробиологии понятия возбудимости. Одно из наиболее точных определений звучит так: возбудимость — это способность некоторых клеток (нервных, мышечных, эндокринных) отвечать на действие стимула закономерными изменениями электрического потенциала своей мембраны. То есть при действии какого-то раздражителя (напомним, что стимул — это вообще-то острый прутик, которым арбакеша побуждает заупрямившегося ослика к началу движения, раздражающий фактор) на мембране этих клеток однотипно, принципиально сходным образом меняются характеристики заряда. А чтобы что-то менять, надо это иметь. Стало быть, возбудимые клетки изначально, ещё до нанесения стимула, обладают зарядом мембраны. Попробуем разобраться, откуда же он берётся и как и почему он может меняться. 1. МОРФО-ФУНКЦИОНАЛЬНАЯ ХАРАКТЕРИСТИКА МЕМБРАНЫ ВОЗБУДИМЫХ КЛЕТОК Для ответа на поставленный только что вопрос нам придётся вспомнить некоторые данные из анатомии, гистологии, цитологии, химии (биохимии в том числе) и физики. Привычнее всего обратиться к нейрону, как функциональной единице нашего мозга. Вспомним, что в нейроне различают сому (или тело клетки), дендриты — как правило (но далеко не всегда!), многочисленные обильно ветвящиеся относительно короткие отростки, и аксоны — как правило (тоже не всегда), относительно длинные маловетвящиеся одиночные отростки, заканчивающиеся пресинаптическими окончаниями, контактирующими со следующими возбудимыми клетками (рис. 2). В центральной нервной системе человека (ЦНС) обнаружено 1011 — 1012 нейронов. Если учесть, что каждый нейрон в среднем принимает по 1000 синаптических входов (а у некоторых количество таких входов более десяти тысяч), можно представить себе сверхсложность нашей информационной системы. Кроме того, сегодня доказано участие в обработке информации сопутствующих нейронам глиальных клеток, что ещё больше расширяет возможности мозга. Но вернёмся к нейронам. Как и положено всякой добропорядочной клетке, нейрон имеет ядро, цитоплазму, митохондрии, комплекс Гольджи, рибосомы и другие органоиды. Наружная поверхность нейрона образована мембраной. Именно она и явится для нас сейчас предметом особого внимания. Рис. 2. Изображен весьма правдоподобный нейрональный модуль, состоящий из трёх нейронов. Хорошо видны тела (сомы) этих трёх нервных клеток с ядром внутри каждой, относительно короткие ветвящиеся дендриты и сравнительно длинные аксоны. Обратите особое внимание на два синапса, которыми верхний нейрон связан с двумя нижними клетками: с дендритом левой (аксо-дендритный синапс) и сомой правой клетки (аксо-соматический) (Эта иллюстрация, а также рис. 19, 50 и 56 скопированы из презентации к публичной лекции К.В.Анохина, 2006). Как известно, клеточная мембрана состоит из двойного слоя липидов (в основном, фосфолипидов), в который встроены различные по структуре, свойствам и функциям белковые молекулы (рис. 3, 4). Рис. 3. Схематическая реконструкция клеточной мембраны. В хорошо структурированный липидный бислой встроены белковые молекулы, обладающие специфическим строением и специфическими функциями. Рис. 4. Биологоправдоподобное изображение клеточной мембраны. Хорошо видны различные белковые глобулы (в том числе ионные каналы), «плавающие» в двойном слое липидных молекул (Эта иллюстрация, а также рис. 5, 10, 12 – 14, 16, 43 и 52 скопированы из книги Николлса и др. «От нейрона к мозгу», 2003). Устойчивость липидного бислоя обеспечивается своеобразными свойствами липидных молекул — их полярным отношением к водной среде. Фосфоглицериновая «головка» липидной молекулы гидрофильна, то есть водорастворима (для проверки можете сходить в аптеку и купить водный раствор глицерина), тогда как образованные жирными кислотами «хвосты» гидрофобны, то есть воду категорически не любят и всячески стараются держаться от неё подальше (опять же можете сами попробовать сотворить водный раствор подсолнечного масла, только продуктивнее это время потратить на хорошую книжку). Благодаря этому свойству липидные молекулы в водной среде (а в нашем организме среда-то как раз водная, ведь и цитоплазма, и межклеточная жидкость — это, по сути, водные растворы разных органических и неорганических соединений) стремятся образовывать компактные скопления, мицеллы, в которых гидрофобные полюса молекул обращены друг к другу — подальше от воды, — а гидрофильные «головки» вполне закономерно развёрнуты наружу, взаимодействуя с водой.Бислойная мембрана — один из примеров такого компактного скопления липидов, очень стабильного, но одновременно текучего (что тоже важно). В самом деле, простейший аналог такой мембраны — янтарная капелька жира на поверхности горячего бульона: и стабильная, и подвижная. Надеюсь, Вы смогли представить себе эту волнующуюся жировую плёнку на поверхности океана цитоплазмы, а теперь вообразите множество айсбергов, то тут, то там возвышающихся над этой поверхностью. Это белки, а точнее — белковые комплексы. У этих белков разное химическое строение, разные свойства, разные названия, разное место в официальной классификации. Но нам сейчас интересно не это, а то, чем эти белки занимаются на мембране, то есть их функции. Попробуем сгруппировать их именно по функциональному признаку (рис. 5). Рис. 5. Разные по функциональному смыслу белки нейрональной мембраны. Представлены упрощённые химические структуры ионных каналов (А, Е) и мембранных рецепторов (B, C, D). Тогда сразу обнаружится несколько принципиально важных классов белковых молекул (комплексов — повторим ещё раз, потому что, как правило, определённую работу на мембране выполняет не одиночная молекула белка, а молекулярный комплекс, состоящий из нескольких субъединиц) (рис. 6): 1) ионные каналы, 2) ионные насосы, 3) рецепторы, 4) ферменты, 5) структурные белки, и т. д. 1 2 3 4 5 Рис. 6. Схема клеточной мембраны. В обозначенный серой полосой липидный би-слой встроены 5 функциональных типов белков: 1 — каналы, 2 — насосы, 3 — рецепторы, 4 — ферменты, 5 — структурные белки. Ограничимся этим перечнем, он пока достаточен для понимания нами основ биоэлектрогенеза. Что же эти белки делают на мембране? Структурные белки стабилизируют текучий липидный бислой. Трудно предположить, что без них нейрон растечётся по субстрату, подобно часам у Сальвадора Дали, но без белков-стабилизаторов мембраны пока никто не видел. Ферменты, как известно, — это биологические катализаторы. Они снижают энергию активации, то есть обеспечивают существенно облегчённый старт химических реакций в организме. А вот что это будут за реакции, зависит уже от специфики конкретных ферментов. Нас будут интересовать прежде всего те ферменты, которые катализируют реакции, идущие на поверхности мембраны нейрона, в подмембранном пространстве (в цитоплазме) и так называемые трансмембранные реакции («пронизывающие» мембрану). Рецепторы — главные коммуникаторы в межнейронной передаче. Эти белки способны с высоким сродством (то есть очень специфично и в относительно низких концентрациях) связываться с биологически активными веществами (медиаторами, гормонами и т.д.) и, переходя в активированное состояние, изменять физиологическую активность принимающей информацию клетки, в первую очередь — менять электрические характеристики мембраны этой клетки. Ионные каналы и насосы — это транспортные белки, обеспечивающие перенос ионов через мембрану возбудимой клетки. При этом каналы пропускают ионы по их градиенту концентрации; это пассивный транспорт, не требующий энергетических затрат. В противоположность этому насосы переносят ионы против градиента концентрации, затрачивая на эту работу энергию АТФ (универсального клеточного аккумулятора энергии). Такой транспорт является активным. 3. ИОННЫЙ ТРАНСПОРТ: КАНАЛЫ И НАСОСЫ Рассмотрим подробнее функции каналов и насосов, ведь именно их работа обеспечивает формирование и изменение зарядов мембраны. Определяя особенности функционирования этих транспортных белков, мы использовали очень важное понятие: химический градиент ионов. Хочется напомнить, что в растворе молекулы диссоциируют на ионы. Наш организм, на 70% состоящий из воды, не исключение: в нём присутствуют и неорганические ионы (Na+, K+, Ca2+, Cl– и т.д.), и органические (например, белковые анионы). При этом распределение этих ионов по разные стороны мембраны далеко не одинаково. Так, в межклеточном пространстве чрезвычайно высока концентрация катионов Na+ и Ca2+, а также анионов Cl–. В цитоплазме этих ионов, наоборот, очень мало, зато там высоки концентрации катионов K+, а также белковых анионов. Таким образом, мы видим, что мембрана возбудимых (да и не только возбудимых) клеток разделяет две химически неоднородные среды (внутри клетки и снаружи) с существенно разными концентрациями ионов (рис. 7). То есть на мембране можно обнаружить (померить) химический градиент. Рис. 7. Схема иллюстрирует ярко выраженное неравновесие ионов по разные стороны мембраны. Внутри клетки явно выше концентрация K+ и белковых анионов, а снаружи преобладают ионы Na+ и Cl–. А теперь представьте себе простой опыт. Небольшой аквариум разделён пополам непроницаемой перегородкой. Перегородка состоит из двух слоёв: стеклянной пластинки (абсолютно непроницаемой для исследуемых нами веществ) и полимерной плёнки, через микроскопические поры которой могут проходить ионы какого-то одного вида. Наливаем в левую половинку аквариума концентрированный раствор интересующего нас вещества (например, простой поваренной соли, которая в водной среде диссоциирует на ионы: NaCl Na++Cl-), а в правую — такой же раствор, но очень разбавленный (рис. 8). Рис. 8. Выравнивание концентраций вещества в растворах, разделённых полупроницаемой перегородкой. Ионы через каналы (поры) просачиваются по градиенту концентрации. Очевидно, что любые доступные способы измерения покажут наличие существенной разницы концентраций (наличие химического градиента) этого вещества по разные стороны перегородки. Убедившись в этом, выдёргиваем стеклянную перегородку. Остаётся только полупроницаемая полимерная мембрана. Очень скоро мы сможем убедиться, что концентрации по обе стороны мембраны начали выравниваться: ионы по градиенту концентрации начали перебираться из более концентрированной среды в менее концентрированную. Понадобилась для этого дополнительная энергия? Нет, достаточным оказалось предоставить системе возможность самостоятельно решить «проблему социальной справедливости» (всех уравнять!), разблокировав каналы утечки. Именно каналы, ведь через них идёт пассивный транспорт, не требующий дополнительной энергии, по градиенту. А вот теперь представим себе обратную задачу: нам надо вернуть исходное состояние неравновесия концентраций в нашем аквариуме (рис. 9). Понятно, что без затрат энергии это не получится (попробуйте достать воду из колодца, не прилагая усилий!). Именно поэтому перекачка ионов против градиента концентрации — работа уже не каналов, а насосов, и на эту работу расходуется энергия АТФ (между прочим, электронасос накачает Вам ведро воды мигом, но посмотрите на счётчик — сколько энергии Вы сожгли!). Мы коснулись чрезвычайно важных моментов в проблеме биоэлектрогенеза. Проанализируем эти два механизма ионного транспорта детальнее. Во-первых, как устроены ионные каналы? Принятая сегодня модель ионного канала основывается на множестве достаточно убедительных, но всё же косвенных измерений. Хочется ещё раз предостеречь читателей от популярного заблуждения: на самом деле очень многое из того, что мы знаем о микроорганизации нейронов (на молекулярном, мембранном уровнях), является результатом мысленных, технических, компьютерных, математических и т.п. реконструкций объектов по многочисленным физико-химическим измерениям; то есть это гипотетические модели объектов, а вовсе не сами объекты, как может показаться из ряда современных книжек. Рис. 9. Для создания химического градиента требуется приложить энергию. Используя энергию распада АТФ, насосы перекачивают ионы против градиента концентрации. Поэтому используемые повсеместно в нейробиологии словосочетания «селективная пора», «ворота» и т.д. нужно понимать как более или менее удачные метафоры, а не как прямое описание объектов (в данном случае — каналов). Унывать, однако, не вижу оснований: технологическая база исследований развивается очень быстро, и то, что недоступно сегодня, становится рутинной повседневностью завтра. Надеюсь, что скоро сможем убедиться в правильности (или неправильности) модельных реконструкций, в том числе и моделей каналов. Подождём немного? Так вот, согласно принятой сегодня модели ионные каналы — это белковые комплексы, состоящие из нескольких субъединиц — молекул белка (рис. 10). Эти белковые комплексы пронизывают липидную мембрану насквозь, что и позволяет происходить трансмембранному переносу ионов. Рис. 10. Реконструкция ионного канала, встроенного в липидную мембрану. Изображены белковые субъединицы, образующие селективную пору (внутри которой красуется одинокий гидратированный ион) и закрытые в настоящий момент ворота (G — gate = ворота). В канале выделяют два функциональных компонента (рис. 11): селективную пору, обеспечивающую выбор (селекцию) тех ионов, которые этот конкретный канал может пропускать, и так называемые ворота, которые Рис.10 обеспечивают дискретность (прерывность) работы канала (как правило, канал активируется только на очень короткий промежуток времени, всё остальное время он закрыт и ионы не пропускает). Рис. 11. Условная схема, иллюстрирующая работу ионного канала. А — селективная пора, через которую у данного вида канала диффундируют только ионы Na+, тогда как ионы Cl– и K+ сквозь мембрану не проходят. Б — ворота; у данного вида канала в данный конкретный момент времени активационная створка ворот (слева) полуоткрыта, а инактивационная (справа) открыта полностью. Напоминаю, что и селективная пора, и ворота — не более чем относительно удачная метафора. Нет времени (да и необходимости) рассматривать многочисленные гипотезы, объясняющие тонкие электрохимические механизмы функционирования обоих компонентов канала. Гораздо более важным является то, что эти механизмы обеспечивают высокую специфичность работы каналов. Существуют каналы Na+, K+, Ca2+, Cl– и др., то есть благодаря отбору в селективной поре эти каналы избирательно пропускают только один из перечисленных видов ионов, препятствуя транспорту остальных. Необходимо подчеркнуть, что эта селективность, избирательность не является абсолютной: как и положено живой системе, каналы имеют право на ошибку и могут случайным образом пропускать и «неправильные» ионы, однако их доля в общем ионном токе незначительна. Принято различать высокоселективные каналы (например, на мембране аксона — Na+ или K+) и низкоселективные (например, на постсинаптической мембране — Na+— K+ или Cl– — K+). Воротный механизм (или просто ворота) — второй функциональный компонент ионного канала. Естественно, ворота — это весьма вольная метафора, однако она вполне отражает задачу этого механизма: переводить канал из закрытого состояния в открытое и наоборот — снова закрывать его. На самом деле, состояние канала определяется кон-формацией (III структурой) образующих его белковых молекул, а конформация, в свою очередь, зависит от энергетики этих молекул. В принципе любое воздействие, изменяющее энергетическую характеристику белка (электрическое поле, повышение или снижение температуры, сдвиги рН и ионного состава, механическое воздействие, химические взаимодействия и т. д.), может вызывать и изменение его конформации. Однако специфика канальных белков в том, что изменения конформации, приводящие именно к активации («открыванию») канала, возникают только на очень специфичные, эксклюзивные для данного типа каналов воздействия. Так, есть каналы, эффективная активация которых возможна только при изменении электрического поля на мембране — их называют потенциалзависимыми (или, по старинке, электровозбудимыми); есть каналы, открывающиеся только по химическому «приказу» от рецептора — это лигандзависимые каналы (лиганды в нейробиологии — вещества, с высоким сродством связывающиеся с данным конкретным рецептором и изменяющие его функциональное состояние, о лигандах мы поговорим подробно чуть позже; химика это определения приведёт в трепет!); есть удивительные каналы, активирующиеся в ответ на механическое воздействие — нетрудно догадаться, что они называются механозависимыми; наконец, особый тип каналов — поры, у них ворота загадочным образом куда-то потерялись (точнее, их вообще никогда не было), поэтому осталась одна селективная пора, которая открыта всегда, независимо от энергетических проблем (рис. 12, 13). Рис. 12. Наиболее вероятное устройство ионного канала с учётом особенностей строения белковых субъединиц. Это очень упрощённая классификация (по транспортируемым ионам и механизму активации). Так как сегодня обнаружено более 150 видов каналов, существуют и более глубинные принципы объединения каналов в группы, учитывающие целый ряд дополнительных факторов, но мы в эти подробности вдаваться не будем. Рис. 13. Возможный механизм активации потенциалзависимого ионного канала: изменение заряда мембраны (обратите внимание на изменение плюсов и минусов по разные стороны липидной мембраны) меняет конформацию белковых доменов, переводя закрытый канал (А) в открытое состояние (В). Единственное, на что хотелось бы обратить внимание, — это существование не только активационных, но и инактивационных ворот. Вопреки бытующему мнению задача этих ворот состоит вовсе не в закрывании ионного канала, а в переводе его в такое состояние, в котором он определённое время не может активироваться. Подобный механизм есть далеко не у всех каналов, но обнаружен, например, у потенциалзависимых Na+-каналов мембраны аксона. Второй механизм ионного транспорта, который мы обозначили, — активный транспорт с помощью насосов. Мембранных насосов известно много (рис. 14). Они могут иметь разную молекулярную структуру, но функция у них одна и та же: перенос веществ (молекул, ионов, протонов… — не принципиально) против градиента концентрации данного конкретного вещества. Стало быть, такой перенос требует затрат энергии. Как правило, источником энергии является АТФ. Напомним, что аденозинтрифосфат (АТФ) — уникальный по своей безотказности молекулярный аккумулятор энергии. При расщеплении макроэргических (высокоэнергетичных) связей между остатками фосфорной кислоты и соответственно частичном разрушении молекулы АТФ выделяется поразительное для такой субмикроскопической структуры количество энергии, идущей на «хозяйственные нужды» клетки. При этом аккумулятор, естественно, разряжается (АТФ→АДФ), а на его зарядку (восстановление целостности молекулы АТФ) требуется чуть больше энергии, чем было получено при распаде. Откуда берётся эта энергия? По крайней мере, отчасти мы едим для «зарядки» АТФ (отчасти, потому что с едой мы ещё поставляем клеткам стройматериалы, а иногда ещё и получаем удовольствие), а уж дышим и вообще только ради этого. Рис. 14. Разные виды транспортных белков, обеспечивающих активный трансмембранный перенос веществ. (А) — Na+-K+АТФаза, (В) — Na+- Ca2+насос, (С) — Cl-- K+- насос, (D) — моноаминовый транспортер. Но вернёмся к насосам. Попробуем понять принцип их работы на примере так называемой Na+-K+-помпы, которая обеспечивает уже обсуждавшееся нами неравновесие ионов Na+ и K+ по разные стороны мембраны нейрона. Более или менее убедительно показано, что этот насос представляет собой достаточно тяжёлый белок, обладающий, как минимум, двумя активными конформациями, в одной из которых (назовём её X) он имеет повышенное сродство к ионам Na+, а в другой (пусть будет Y) — к ионам K+. На рис. 15 показана очень упрощённая схема работы Na+-K+-помпы. Рис. 15. Условная схема, иллюстрирующая механизм работы ионного насоса. В качестве примера представлена Na+-K+-помпа. Объяснения в тексте. В подмембранном пространстве белок-транспортер в конформации X связывается с ионом Na+ и переносит его на наружную поверхность возбудимой клетки (при этом удаётся «обмануть» градиент Na +, который, как вы помните, направлен внутрь). Там белок расстаётся с Na+ и переходит в конформационное состояние Y. Теперь «на борт» берётся уже ион K + (снова обманут градиент, ведь K+ транспортируется через мембрану в связанном с белком виде — это другое вещество, совсем не K+!), и челнок ныряет внутрь клетки. Здесь — скорое расставание с K+ и возвращение в конформацию X. Круг (цикл) замкнулся. А куда же уходит энергия АТФ? Вспомните, мы говорили о том, что у белков определённому уровню внутренней энергии соответствует определённое конформационное состояние. Отсюда и решение обратной задачи: если конформация изменилась, значит, что-то произошло с энергетикой белковой молекулы, то есть энергия АТФ расходуется на конформационные изменения. И вот здесь очередная демонстрация фантастического совершенства жизни: оказывается, энергетической подпитки требует только переход Y→X (внутри клетки), а обратное изменение конформации (X→Y) происходит самопроизвольно. Объяснение очевидно: состояние Y энергетически более выгодно для данного белка-транспортера (в этом конформационном состоянии у него меньше свободной энергии, он менее возбуждён), чем состояние X. Таким образом, получается, что энергия АТФ тратится только на внутриклеточный переход Y→X. Мало того, работа помпы настолько экономична, что одна молекула АТФ обеспечивает перекачку через мембрану сразу 5 ионов (3+2), то есть 2,5 «оборота» насоса. Этому способствует и так называемая электрогенность насоса, его способность использовать, кроме АТФ, и энергию электрических зарядов, формирующихся на мембране. Если после чтения предыдущей пары страниц Вам всё ещё кажется, что такое объяснение работы насоса слишком уж мудрено, взгляните на рис. 16: идея-то та же, просто графически представлена в другом, уже знакомом Вам виде. Думаю, Вы без труда сами проследите всю цепочку событий. Только ещё раз прошу не забывать, что это всего лишь более или менее правдоподобные модели: не будете же Вы всерьёз думать, что ионы так похожи на шарики для пинг-понга?! Рис. 16. Ещё одна иллюстрация на тему Na+-K+-помпы. Попробуйте самостоятельно пройти по всему циклу A-B-C-D-E-F-A. 4. МЕМБРАННЫЕ ПОТЕНЦИАЛЫ: ПП, ПД, ВПСП, ТПСП, МПКП, ПКП и т. д. Любого здравомыслящего читателя набор аббревиатур, вынесенных в заголовок этой главы, должен привести в чрезвычайно унылое состояние. Просим набраться терпения: очень скоро Вы увидите, что всё не так мрачно, как кажется с первого взгляда. Прежде всего речь в этой главе пойдёт о главной электрической составляющей информационных процессов в нервной системе и в возбудимых тканях вообще. Мы будем отвечать на вопрос, поставленный в конце главы 1: что такое мембранный потенциал и как и почему он может меняться? Напомним, что главным «языком» мозга является язык нервных импульсов, или импульсный код (рис. 17). Рис. 17. Простая имитация импульсной активности нейрона, созданная с помощью всего двух клавиш компьютера. Можно утверждать, что при наличии соответствующей регистрирующей аппаратуры сейчас, в это мгновение, в Вашей голове можно было бы обнаружить нейрон, разряжающийся именно в таком режиме. На рисунке хорошо видно, что после каждого импульса возникает пауза, которая иногда длится мгновение, сопоставимое по длительности с самим импульсом, а иногда растягивается на довольно солидный отрезок времени. Эта пауза называется межимпульсным интервалом и отражает состояние относительного покоя мембраны клетки. Заметим, что чередование импульсов и межимпульсных интервалов (так называемый рисунок паттерна) является одним из возможных кодов нейрона, хотя его роль в обработке информации, судя по всему, весьма невелика. Для нас же сейчас важно выяснить, каким электрическим характеристикам мембраны соответствует состояние её покоя и активности (в момент генерации импульса). 4.1. Потенциал покоя (ПП) мембраны и причины его возникновения Постарайтесь представить себе достаточно несложный по сегодняшним возможностям эксперимент. Прежде всего, что Вы будете делать, чтобы померить напряжение в сети, от которой питается Ваша настольная лампа? Думается, многим из Вас знаком прибор под названием тестер (официально он называется омампервольтметр или как-то похуже), в котором есть непосредственно измеритель (в простейшем варианте струнный гальванометр) и два провода — электрода, кончики которых мы можем аккуратно ввести в электрическую розетку. Обнаружив, что измеритель показал 205—210 В (на 220 рассчитывать не очень приходится, учитывая, что в нашей стране всё воруют), мы сразу успокаиваемся: напряжение измерено. А теперь представьте себе, что Вам надо померить напряжение (давайте начнём пользоваться более «физиологичным» понятием — потенциал) на мембране нейрона. Принцип-то измерения будет точно таким же: нужен измеритель и нужны электроды. Только надо учесть миниатюрность всего того, что мы собрались измерять, поэтому и электроды будут особо тонкими (микроэлектроды), и сам измеритель гораздо чувствительнее, чем для бытовых нужд. Ведь в нейронах (забегаем вперёд) потенциалы на 3 порядка слабее, чем в розетке: милливольты (мВ), а не вольты. Как раз теперь самое время поговорить о методах измерений электрических процессов в живых системах. В принципе не так уж и важно, что мы меряем: потенциалы отдельных нейронов (или даже участков их мембраны), их больших или сравнительно небольших скоплений — модулей (так называемые фокальные потенциалы), суммарные потенциалы больших участков мозга (например, ЭЭГ — электроэнцефалография), потенциалы сердца (ЭКГ — электрокардиография), или что-то другое. Подход к измерениям достаточно стандартен (рис. 18). Рис. 18. Принципиально необходимые компоненты электрофизиологической аппаратуры (объяснения в тексте). современной Нужны электроды для отведения потенциала от объекта измерения. Если объект маленький, используются микроэлектроды (как правило, стеклянная микропипетка, заполненная каким-то электролитом, например, KCl), если покрупнее — макроэлектроды. Так как потенциалы и токи в живых системах очень слабы, необходимо уметь их усиливать при регистрации. Поэтому во всей электрофизиологической аппаратуре есть специальные усилители. При усилении сигнала возникает серьёзная проблема: вокруг нас очень много всяческих электрических и радиотехнических приборов (радиотелефоны, телевизоры, пылесосы, трамваи и прочие достижения цивилизации), создающих массу помех; «шумят» и живые клетки. Усиливая интересующий нас сигнал, мы автоматически усиливаем и сопровождающие его шумы. И здесь есть опасность вообще потерять сигнал в этом море помех. Поэтому в современных электрофизиологических приборах обязательно присутствуют специальные фильтры, позволяющие вырезать сигнал из шума. И, наконец, зарегистрированный, усиленный и очищенный от помех сигнал надо как-то увидеть и зафиксировать, то есть сохранить. Так как сегодня аппаратура для измерений компьютерная, с этим проблем нет. Надо только преобразовать те электрические изменения, которые мы поймали нашими электродами (так называемый аналоговый сигнал), в цифровую форму, понятную компьютеру. Это умеют проделывать АЦП — аналоговоцифровые преобразователи (есть, кстати, и обратные преобразователи — ЦАП). Теперь уже ничто не помешает увидеть зарегистрированные биопотенциалы на экране монитора компьютера, записать их в память, обработать любым способом и т. д. Однако вернёмся к нашему эксперименту. Представьте себе, что мы получили возможность поработать с отдельным нейроном. Не слишком важно, что это за нейрон, и каким конкретным методом мы будем к нему подбираться. Это может быть нервная клетка одного из отделов мозга (рис. 19), до которой мы добрались, предварительно занаркотизировав (подобно операционному наркозу в хирургических клиниках) животное, закрепив его голову в специальном станке (стереотаксисе), сняв участок скальпа, проведя трепанацию черепа (проще говоря, просверлив череп в строго определённом месте) и с по мощью микроманипулятора введя микроэлектрод в чётко известную структуру мозга (вводить мы его будем по координатной сетке трёхмерного атласа — такие атласы есть практически для всех лабораторных животных). Это может быть один из нейронов тончайшего переживающего среза одной из мозговых структур (переживающими такие срезы — слайсы = slices — называют не за особую эмоциональность, а за то, что в искусственно созданной среде эти мозговые срезы надолго переживают своего хозяина — крыску, хомячка, морскую свинку и т. д.). Это может быть одна из клеток культуры нервной ткани: для тех, кто наслышан про пресловутое клонирование, возможность выращивания в чашке Петри целой популяции одинаковых клеток из одной первоначально взятой не кажется фантастикой. Наконец, это может быть периферический отросток (лучше —аксон) нервной клетки какого-нибудь животного; первые эксперименты, подобные обсуждаемому нами, так и делались: в 40—50-е годы ХХ века великолепная группа физиологов под руководством Алана Ходжкина смогла понять механизмы возникновения биопотенциалов благодаря работе на гигантском аксоне кальмара (сами-то кальмары не были гигантскими, просто для эффективной работы реактивного двигателя этого животного — колокола, из-под которого «выстреливает» вода, — нужна очень высокая скорость распространения нервных импульсов вдоль колокола, а чем больше диаметр волокна, тем быстрее по нему бегут импульсы, поэтому у знакомых всем по салатам кальмаров аксоны мотонейронов можно увидеть невооружённым глазом). Рис. 19. Вполне правдоподобный рисунок, воспроизводящий принцип микроэлектродного отведения потенциалов от одиночного нейрона. Итак, наша первая насущная задача — выяснить, есть ли на мембране аксона, находящегося в состоянии покоя (во время межимпульсного интервала), какой-либо электрический потенциал, и если есть, то какой. Используем уже упоминавшийся микроэлектрод. Подводим его кончик непосредственно к мембране аксона. Второй электрод — здесь вполне подойдёт и макроэлектрод — помещаем где-то поблизости, не очень заботясь о точности его расположения: так как нам нужно мерить разность потенциалов, обязательно нужны два электрода — активный (микроэлектрод) и индифферентный (можно макро-). Подсоединяем оба электрода к измерительной аппаратуре, включаем и… на нашем мониторе мы видим тоскливую прямую: разность потенциалов (давайте говорить проще — потенциал) равна нулю, то есть её просто нет при расположении обоих электродов снаружи от мембраны (рис. 20). Теперь начинаем осторожно, с помощью микроманипулятора, опускать микроэлектрод. В какой-то момент его кончик проткнёт мембрану и войдёт внутрь нервной клетки (не надо пугаться: ведь мембрана — это жидкий бислой липидов, так что с нейроном ничего не случится; вспомните: «нож сквозь масло»). И именно в этот момент мы увидим на экране резкий скачок уровня потенциала — до 60—70 мВ, причём, судя по показателям, внутренняя поверхность мембраны окажется заряженной отрицательно по отношению к наружной поверхности. мВ Рис. 20. Измерения мембранного потенциала нейрона в состоянии покоя, то есть во время межимпульсного интервала. Пока микроэлектрод находится на поверхности мембраны, регистрируется отсутствие значимой разности потенциалов между ним и макроэлектородом, который может находиться достаточно далеко от данного нейрона. Как только с помощью микроманипулятора мы опускаем кончик микроэлектрода и «входим» под мембрану (этот момент обозначен стрелкой), наша измерительная аппаратура улавливает резкий скачок потенциала: внутренняя поверхность мембраны оказывается значительно (на десятки милливольт) более электроотрицательной, чем наружная. Это и есть потенциал покоя мембраны (ПП). Итак, установлен первый вполне конкретный научный факт: во время межимпульсной паузы, в состоянии релаксации, покоя, внутренняя поверхность мембраны нейрона заряжена отрицательно относительно наружной, и уровень этого потенциала колеблется (в зависимости от условий конкретного опыта) около 60—70 мВ. Поздравляю: мы зарегистрировали ПП — потенциал покоя мембраны! На конкретные значения ПП не стоит обращать особого внимания: в зависимости от того, какую возбудимую клетку мы обследуем, у какого животного, в каких условиях, значения могут несколько меняться. Здесь важно другое: устойчивый, всегда воспроизводимый факт наличия отрицательного электрического заряда на внутренней поверхности мембраны. Откуда же он взялся? Для ответа на этот вопрос вспомним недавний разговор про мембрану и её свойства. И снова речь пойдёт об особенностях химического градиента ионов по разные стороны мембраны и об ионных каналах. В своих, ставших сегодня уже классическими экспериментах сотрудники группы Ходжкина обнаружили, что в состоянии покоя на нейрональной мембране открыты каналы одного-единственного вида: + K -поры. Давайте ещё раз вспомним, что означает такое название: во-первых, это поры, стало быть, ворота у них отсутствуют, и они открыты всегда; вовторых, это K+-каналы, то есть они пропускают почти исключительно ионы K+ по градиенту концентрации. А как направлен химический градиент K +? Изнутри наружу клетки. То есть ионы K+ будут выходить через эти каналы наружу (рис. 21). А теперь самый ключевой вопрос: до каких пор будет происходить этот выход K+? Рис. 21. В состоянии покоя ионы K+ через поры выходят из клетки по градиенту концентрации. Ответ, казалось бы, напрашивается сам собой: до тех пор, пока концентрация этих ионов по разные стороны мембраны не станет одинаковой. И этот ответ был бы верным (ведь он вполне в русле термодинамических законов), если бы не одно существенное обстоятельство: мы имеем дело не с простыми электрически нейтральными веществами, а с ионами. А в этом случае устранение одной «несправедливости» (неравенства концентраций) неизбежно приведёт к возникновению другой «несправедливости» — неравенству электрических зарядов по разные стороны мембраны. Ведь с каждым выходящим из клетки по градиенту концентрации ионом K+ клетка одновременно теряет и положительные заряды, которые перетекают на наружную поверхность мембраны (рис. 22). Рис. 22. Выход ионов K+ из клетки по градиенту концентрации приводит к созданию электрического градиента на мембране: внутри клетки накапливаются «осиротевшие» белковые анионы (следовательно, отрицательные заряды), а снаружи катионы K+, то есть положительные заряды. То есть в клетке формируется дефицит плюсов (зато избыток минусов — не забывайте, например, про белковые анионы, которым через каналы не пройти при всём желании), а вокруг неё — избыток тех же самых плюсов. Стало быть, возникает электрическое неравновесие, электрический градиент на мембране Вернёмся к ключевому вопросу: до каких же пор возможен выход ионов + K из клетки? Если до сих пор не догадались, попробуем вспомнить младшешкольную физику. Мы с Вами стараемся понять механизмы, формирующие ПП мембраны. При каких условиях некое вполне материальное тело находится в состоянии покоя (или равномерного прямолинейного движения — что, по существу, одно и то же)? Представьте себе, что на Вашем письменном столе лежит книжка. Почему она не падает на пол и не взлетает к потолку? Надеюсь, Вы вспомнили: потому что силы, действующие на эту книжку, равны и направлены в противоположные стороны (рис. 23). Сила тяжести тянет эту книжку вниз, а сила сопротивления (противодействия) опоры (стола) столь же решительно препятствует этому. Рис. 23. Книжка будет спокойно лежать на столе при условии равновесия двух диаметрально противоположно направленных сил: силы тяжести (Fтяж) и силы сопротивления опоры (Fсопр). Какое это отношение имеет к ионным процессам на мембране нейрона? Самое прямое! Мы с Вами стараемся понять механизмы, формирующие потенциал покоя (ПП) мембраны. Мы (вслед за группой Ходжкина) выяснили, что в покое на мембране формируются две силы: химический градиент ионов K+ (провоцирующий выход этих ионов из клетки через поры) и встречный электрический градиент (то есть потенциал), связанный с потерей клеткой положительных зарядов. Тогда становится понятным, что условием равновесного («спокойного») состояния мембраны является равновесие этих двух диаметрально противоположно направленных сил (рис. 24). Необходимо подчеркнуть два момента: во-первых, когда мы говорим о выходе ионов калия из клетки и «выстраивании» их по наружной поверхности мембраны, речь идёт исключительно о тончайшем примембранном слое, чуть дальше в межклеточном пространстве концентрация K+ по-прежнему будет значительно ниже, чем внутри клетки. Во-вторых, описанное нами на качественном уровне объяснение механизма формирования ПП может быть представлено гораздо строже, если мы воспользуемся так называемым уравнением Нернста. Рис. 24. Потенциал покоя (ПП) или равновесный калиевый потенциал (EK), существующий на мембране возбудимой клетки в период межимпульсного интервала, уравновешивает химический градиент по калию ([K+нар] / [K+внутр]). Вопреки прозрачным намёкам некоторых учебников немецкий учёный Нернст не был физиологом (тем более — нейрофизиологом, ведь в XIX веке эта наука только зарождалась); он был выдающимся физхимиком. И своё уравнение диффузии ионов через полупроницаемые мембраны он вывел как общую закономерность, а вовсе не для описания транспорта K + через нейрональную мембрану. Но знаменитым стало именно нейробиологическое приложение его закона. Итак, что же это за формула? EK = (R T / F Z) ln ([K+нар] / [K+внутр]), где EK — электрический потенциал мембраны по K+, R — газовая постоянная, T — абсолютная температура, F — число Фарадея, Z — валентность иона K+, [K+нар] / [K+внутр] — градиент концентрации ионов K+ снаружи и внутри клетки. Учитывая, что эта книга написана в том числе и для убеждённых гуманитариев, постараемся помочь им преодолеть священный ужас перед любыми формулами. Вглядитесь повнимательней: первые четыре переменные в данном уравнении — это вполне конкретные численные значения, которые легко можно найти в любом физическом или химическом справочнике. Логарифмирование — не более чем математический приём. Что же тогда остаётся? То, что равновесный калиевый электрический потенциал на мембране нейрона при стандартных условиях зависит от соотношения концентрации ионов K+ снаружи мембраны к их концентрации внутри: EK – ([K+нар] / [K+внутр]). Значком мы рискнули обозначить некое принципиальное соответствие правой и левой частей выражения, потому что знак равенства ставить теперь мы уже не имеем права; а минус перед правой частью выражения неизбежно должен был появиться при уходе от логарифмирования (вспомните: при логарифмировании дроби, в которой числитель меньше знаменателя, получается отрицательное число). Таким образом, при анализе уравнения Нернста смысловое содержание нашего качественного рассуждения вполне подтвердилось. Хочется ещё раз подчеркнуть, что формализация электрохимического сопряжения приложима ко всем вариантам ионных токов через мембраны. Не случайно на базе полного уравнения Нернста (в него входят ионные градиенты не только для калия, но и для натрия и хлора) Гольдманом, а чуть позднее Ходжкиным, Хаксли и Катцем были прописаны уравнения «постоянного поля» нейрона, ставшие сегодня одной из основ математического моделирования электрических процессов на мембране нейронов (заметим в скобках, что к передаче информации подобного рода модели имеют самое прямое, но — увы! — пока ещё весьма далёкое отношение). Если у Вас сложилось впечатление, что с причинами возникновения ПП теперь всё ясно, вынужден Вас огорчить. Оказывается, если действительно подставить в уравнение Нернста все необходимые численные значения, то вычисленный уровень ПП окажется не совсем таким, как экспериментально измеренный. Несложные вычисления приведут к значениям EK –85-90 мВ, а реальные измерения дают, как Вы помните, цифры около –60—70 мВ. Почему такое расхождение, нет ли ошибки в самой гипотезе? И да, и нет. Нет, потому что бесчисленные измерения абсолютно однозначно показывают, что именно калиевый ток через мембрану является главным, доминирующим фактором формирования ПП. Да, потому что мы имеем дело с живой системой, в которой не может быть ничего абсолютного. Сегодня хорошо известно, что в покое мембрана нейрона пропускает по градиентам концентраций не только ионы калия, но и натрий, хлор и т. д. Другое дело, что их суммарный вклад в конечный уровень ПП невелик, хотя и, как Вы видите, заметен. 4.2. Потенциал действия (ПД), или нервный импульс (спайк) Теперь, когда мы выяснили, что в покое мембрана нейрона уже заряжена (поляризована), пришла пора обратиться непосредственно к потенциалу действия (ПД). Представьте, что у регистрируемого нами нейрона закончился межимпульсный интервал и мы дождались импульса. Или, чтобы не ждать милостей от нейрона, мы через так называемые стимулирующие электроды (к замечательному прибору электростимулятору мы ещё вернёмся попозже) нанесли на поверхность этого нейрона слабенький (всего-то несколько милливольт) стимул. И вспомните рис. 17. На нём отдельные импульсы выглядят как вертикальные чёрточки, что неудивительно: длительность ПД составляет одну-две миллисекунды (мс) — интервал времени, уловить который без аппаратуры человеку не дано. Да и на слух, если нашу регистрирующую аппаратуру дополнить динамиками, импульсная активность нейрона звучит как беспорядочная автоматная стрельба на бейрутских улицах. Что же делать, если изучаемый нами объект так миниатюрен и по амплитуде (десятки милливольт), и по длительности (миллисекунды)? Ответ, вообще говоря, хорошо известен: чтобы увидеть инфузорию-туфельку, надо взять микроскоп. Нам даже и микроскопа не надо, чтобы увеличить наш ПД. Надо просто растянуть на экране компьютера ось времени (X) и ось амплитуды ПД (Y), и мы увидим нервный импульс во всех деталях (рис. 25). Оказывается, ПД имеет достаточно сложную форму, и, двигаясь по оси времени, мы можем разложить его на 5 фаз: фаза деполяризации — от исходного уровня ПП до 0 (то есть мембрана в это мгновение электронейтральна); реверсия ПД или овершут — уровень ПД выше 0 (то есть в этот момент мембрана внутри становится заряженной положительно, а не отрицательно, как было в покое); фаза реполяризации — уровень мембранного потенциала демонстрирует явственное стремление вернуться к исходному уровню ПП; следовая деполяризация — спад графика ПД становится менее крутым, то есть к исходному уровню ПП потенциал мембраны возвращается несколько медленнее, чем во время реполяризации; следовая гиперполяризация — в этот короткий период времени мембрана поляризована даже больше, чем в состоянии покоя. Не стоит сразу пугаться этих новых терминов: они вполне доступны пониманию и весьма информативны. Мы теперь знаем, что в покое мембрана заряжена, или поляризована. Это и есть коренное слово. А дальше — работа с приставками: де- — приставка негативная, отрицательная; действительно, и в 1-й, и в 4-й фазах мембрана менее поляризована, чем в покое; ре- — возвратная приставка (ретро-), соответственно, в 3-й фазе мы видим возвращение ПД (вектор) к исходному уровню. Гипер- — слишком, чересчур; действительно, в 5-й фазе мембрана поляризована слишком сильно, больше, чем в покое. Наконец, обозначения 2-й фазы выходят за рамки этого правила, просто и реверсия, и овершут — это переворачивание чего-то, в нашем случае — мембранного потенциала. Е, мВ t, мс Рис. 25. Потенциал действия (ПД) мембраны нейрона или нервный импульс. Цифрами обозначены фазы ПД: 1 — деполяризация; 2а и 2б — реверсия или овершут; 3 — реполяризация; 4 — следовая деполяризация; 5 — следовая гиперполяризация. Но назвать ещё не значит объяснить. Давайте разбираться, почему мембрана нейрона в ответ на электрический стимул так прихотливо меняет свой заряд. И снова пойдём по стопам группы Ходжкина. Начнём с фазы деполяризации. Почему в ответ на внешний стимул поляризация мембраны начинает очень быстро уменьшаться? Оказывается, в это мгновение на мембране начинают активироваться потенциалзависимые натриевые каналы (рис. 26). Рис. 26. В ответ на электрический стимул на мембране нейрона наряду с постоянно открытыми K+-порами открываются потенциалзависимые Na+каналы, обеспечивающие входящий натриевый ток. В названии этих каналов — 2 ключевых слова: во-первых, они селективны по Na+; а во-вторых, они активируются только при вполне определённых изменениях электрического потенциала мембраны. И вот этот второй момент заставляет нас вернуться к понятиям «стимул» и «электростимулятор». Конечно, можно во время исследования спокойно ждать, когда на нейроне возникнет очередной нервный импульс, тем более что всегда молчащих нейронов по определению не бывает. Но гораздо удобнее, если мы будем точно знать, когда возникнет ПД, а для этого проще всего самим спровоцировать его возникновение. это можно сделать, нанеся на нейрон адекватный (соответствующий его свойствам) стимул. Раз речь идёт о потенциалзависимых каналах мембраны, то стимул должен быть электрическим. И сформировать этот стимул мы можем с помощью лабораторного электростимулятора (рис. 27). Хорошо известно, что электрические процессы в наших нейронах нужно имитировать не привычным для нас переменным током, а импульсами постоянного тока. Уже совсем скоро (при рассмотрении синапса) мы с Вами поймём, с чем это связано. Именно для имитации реально происходящих в нейронах процессов и были сконструированы первые электростимуляторы данного типа. Итак, мы возвращаемся к ПД. Что же происходит в нейроне сразу в ответ на стимул? Если мы подадим на мембрану слабый (всего несколько милливольт) прямоугольный катодический (то есть идущий от отрицательного полюса нашего стимулятора) стимул, возникнет начальная деполяризация мембраны. Этого незначительного сдвига мембранного потенциала в сторону 0 достаточно, чтобы активировалась первая порция потенциалзависимых Na +каналов. Рис. 27. Примитивный лабораторный электростимулятор позволяет регулировать амплитуду стимула (от 0 до нескольких десятков тысяч милливольт), его длительность (от долей миллисекунды до десятков секунд) и полярность (+ или –). Стимул представляет собой прямоугольный (Побразный) импульс постоянного тока. Естественно, через них с существенно большей вероятностью, чем в покое, возникнет входящий Na+-ток. Катионы Na+, входя в клетку по градиенту концентрации, уменьшают электроотрицательность внутренней поверхности мембраны, то есть деполяризуют её ещё больше (рис. 28). Рис. 28. Цикл Ходжкина, обеспечивающий восходящую фазу (фазу деполяризации) ПД. Обратите внимание: стимул нужен только на начальном этапе — для запуска цикла Ходжкина, а дальше процесс развивается самопроизвольно, по механизму положительной обратной связи. В результате возникает лавинообразно нарастающий входящий ток катионов Na+. Круг замкнулся, и теперь все будет повторяться и повторяться. Впервые этот механизм был описан всё той же группой Ходжкина и в мировой литературе получил наименование «цикл Ходжкина». По существу, это классический пример системы с положительной обратной связью. Здесь каждый предыдущий шаг подстёгивает следующий за ним, усиливая эффект. В биологических системах такого рода механизмы встречаются не слишком часто (в тех случаях, когда или требуется усилить эффект, или нужно синхронизировать изначально независимые процессы — например, при брачном поведении у животных). В технике системы с положительной обратной связью мы видим сплошь и рядом: это один из самых распространённых видов усилителей. Цикл Ходжкина поучителен, по крайней мере, по двум причинам. Во-первых, мы видим, как остроумно природа решила проблему быстрого усиления входящего тока Na+ и, соответственно, деполяризации мембраны. Не случайно во многих книжках процесс нарастания входящего натриевого тока называют лавинообразным. Во-вторых, становится более понятной роль начального стимула в запуске ПД. Это, по существу, аналог стартёра в современном автомобиле: маленький слабый электромоторчик, работающий от аккумулятора, может раскрутить тяжёлый на подъём двигатель внутреннего сгорания («завести» его), а дальше он будет работать вполне самостоятельно («самопроизвольно», как сказали бы мы). На стартёре, в принципе, тоже можно проехать несколько метров, но лучше этого не делать из гуманистических соображений. И на мембране нейрона стимул тоже нужен только «на старте» ПД, а дальше процесс поддерживает себя сам. Очень важный вопрос «а всякий ли стимул запустит ПД?» мы проанализируем немножко попозже. И ещё один вопрос, связанный с восходящей фазой ПД. Если цикл Ходжкина — самоусиливающийся процесс, то почему он не развивается бесконечно, что служит ограничителем входящего тока Na +? Ответов несколько. Прежде всего это уже обсуждавшееся нами уравнение Нернста, описывающее взаимосвязь электрического потенциала полупроницаемой мембраны и концентрационных градиентов. Естественно, что изменение градиента (одной из действующих сил) неизбежно ведёт и к изменениям потенциала (второй силы), и далее по циклу. Число потенциалзависимых каналов на данном участке мембраны хоть и велико, но не бесконечно. А если учесть, что только что активировавшийся канал не может сразу же открыться вторично (о причинах этого мы поговорим чуть позже), становится понятным наличие ограничений на циклические процессы на мембране. Есть и ещё несколько менее значимых факторов, обсуждение которых мы вынесем за скобки. Итак, вопрос о механизме деполяризации более или менее прояснился. Но ПД развивается дальше, и после пика реверсии, когда внутренняя поверхность мембраны становится положительно заряженной, наступает фаза реполяризации. Какие мембранные механизмы обеспечивают её развитие? Мы должны назвать, как минимум, три фактора: в результате инактивационных процессов закрываются Na+-каналы и прекращается (точнее, минимизируется) входящий натриевый ток; продолжается выходящий ток K+ через поры; развивается дополнительный выходящий ток K+ через потенциалзависимые калиевые каналы (рис. 29). Рис. 29. Фаза реполяризации ПД обусловлена, как минимум, тремя факторами: а) прекращением входящего тока Na+; б) продолжением выхода K+ через поры;в) дополнительным выходящим током K + через потенциалзависимые калиевые каналы. Думается, Вы почувствовали, что если два первых фактора вполне очевидны, то третий требует дополнительных разъяснений. В самом деле, что же это за «дополнительные» потенциалзависимые K +-каналы, которые активируются так своевременно? И этому открытию уже полвека. Действительно, мы же нигде не оговаривали, что эксклюзивным правом открываться в ответ на изменения потенциала мембраны обладают только Na+-каналы. Белковые субъединицы, образующие K+-каналы, тоже могут быть чувствительными к этим изменениям. Тогда, может быть, для их активации требуются какие-то другие, отличающиеся от необходимых для Na+-каналов параметры изменений потенциала мембраны? Да нет, диапазон примерно тот же. А раз так, значит, эти K +-каналы активируются в ответ на тот же начальный стимул, что и Na+? Ответ пугающе прост: да! Вдумайтесь в этот ответ! Если каналы для двух катионов (натрия и калия) открываются в ответ на сходные изменения потенциала мембраны, то есть на один и тот же стимул, а градиенты концентраций этих катионов направлены навстречу друг другу, то должен возникнуть встречный ток двух этих ионов, и об изменениях потенциала на мембране можно вообще забыть. Но эти изменения есть, иначе информация Вашим мозгом не могла бы обрабатываться, и Вы не могли бы читать эти строки. Где же нестыковка? Какие факторы не были учтены? Чтобы разобраться, нам придётся ввести одно из ключевых понятий мембранологии: проницаемость мембраны для ионов (g). Этот термин означает, сколько данных ионов (Na+, K+, Cl– и т.д.) пропустит единица площади данной мембраны (например, 1 квадратный нанометр) в единицу времени (например, в 1 миллисекунду). Когда мы упоминали уравнения Ходжкина — Хаксли, Катца и др., вскользь было сказано, что они являются развитием идеи Нернста; так вот, одним из элементов этого развития как раз и было введение в уравнения показателя проницаемости мембраны для данных конкретных ионов. Нам этот показатель сейчас нужен для того, чтобы разобраться в загадке «встречных перевозок» катионов через мембрану. Оказывается, хотя процесс активации ионных каналов Na+ и K+ и начинается одновременно (в ответ на стимул), скорость нарастания этой активации, и соответственно проницаемости, существенно различна для ионов Na + и K+ (рис. 30). Скорость нарастания (а затем и спада) натриевой проницаемости намного выше, чем калиевой. Поэтому массированный вброс Na+ в первые мгновения ПД существенно опережает медленно нарастающий выход K+, даже несмотря на то, что калиевая проницаемость мембраны изначально больше (за счёт постоянно открытых калиевых пор). Соответственно, входящий и выходящий катионные токи разведены по времени, и вслед за деполяризацией развивается реполяризация мембраны. Рис. 30. В одном временном масштабе показаны ПД (А) и изменения проницаемости мембраны (Б) для ионов Na+ и K+. Хорошо видно, что проницаемость для Na+ резко возрастает в ответ на стимул и так же быстро падает почти до нуля. Проницаемость для K + начинает возрастать одновременно с натриевой, но увеличивается и спадает гораздо медленнее. Кроме того, проницаемость мембраны для K + уже изначально гораздо выше, чем для Na+, так как калиевые поры остаются открытыми всё время. По существу, с завершением фазы реполяризации завершаются и главные информационно значимые процессы на мембране. Далее, как Вы помните, развиваются следовые процессы, характер которых может различаться в разных возбудимых клетках (вплоть до перемены местами следовой деполяризации и гиперполяризации), и которые, видимо, «не считываются» нашим белковым компьютером напрямую. Но это не значит, что они не влияют на мозговые коды и соответственно на обработку информации. Сегодня существуют вполне биологоправдоподобные модели нейронных сетей, в которых главным изменяемым параметром является период следовой гиперполяризации. Эти виртуальные «мозги» вполне успешно справляются с весьма сложными задачами, что подтверждает небезразличность для информационных процессов следовых («остаточных», как может казаться) изменений поляризации. Однако эти тонкости выходят за пределы нашей книжки. Нам хотелось бы показать на примере следовых процессов значимость ещё одного транспортного механизма мембраны. Речь пойдёт об ионных насосах. Мы с Вами уже рассматривали общие принципы их работы, так что теперь можем обсудить только самое главное. Посмотрите ещё раз на рисунок ПД (рис. 31). Совершенно ясно, что к моменту завершения процесса реполяризации соотношения ионных градиентов по обе стороны мембраны явно не соответствуют «нормативным параметрам», характерным для ПП. Рис. 31. И снова ПД со всеми его фазами. Обратите внимание на точки I, II и III. Как видите, они лежат на одной прямой, то есть уровень мембранного потенциала в них один и тот же (как мы договорились, около –70 мВ). А что Вы скажете об ионных градиентах по обе стороны мембраны? Ведь совершенно очевидно, что в I и III точках они совпадают или почти совпадают. А во II точке? Тем не менее, после окончания следовых процессов мембрана вполне готова к генерации нового импульса (строго говоря, она готова к этому и несколько раньше, но об этом мы ещё поговорим), потому что к этому моменту уровни катионных градиентов успевают вернуться к исходным значениям. Единственным механизмом, способным так быстро восстанавливать ионные градиенты, является ионный насос. В разбираемом нами случае это Na+-K+-ионообменник, или Na+-K+-помпа. Вы, конечно, помните, что такой насос качает ионы против градиента концентрации, затрачивая на это энергию АТФ. Вопреки декларациям многих современных учебников я утверждаю (основываясь на экспериментальных данных, естественно), что эти насосы работают непрерывно, постоянно поддерживая неравновесие концентраций ионов по разные стороны мембраны. Как Вы уже поняли, во время ПД такое неравновесие нарушается особенно сильно. Замечательно, что натрийкалиевый насос в это время резко увеличивает свою активность, чему способствует его особое свойство — электрогенность (это значит, что он активируется при изменении мембранного потенциала, то есть именно во время ПД). А дальше — своеобразный эффект маскировки: пока потенциалзависимые каналы открыты и пропускают ионы по их концентрационным градиентам, интенсивная работа насосов относительно незаметна и обнаруживается только при достаточно тонкой постановке экспериментов, когда же процессы деполяризации и реполяризации остались позади — активный ионный транспорт с помощью насосов проявляется во всей красе. Таким образом, именно во время следовых процессов мы наиболее явно обнаруживаем присутствие на мембране энергозависимых ионообменников — насосов (рис. 32). Рис. 32. Относительно полная схема мембраны нейрона, описывающая базовые процессы, которые обеспечивают формирование ПП и ПД: K +-поры (открыты всегда и участвуют как в ПП, так и в ПД), Na+-потенциалзависимые каналы (главный фактор фазы деполяризации ПД), K +-потенциалзависимые каналы (наряду с порами обеспечивают фазу реполяризации ПД), Na +-K+помпа (восстанавливает и поддерживает катионные градиенты на мембране). 4.3. Свойства потенциала действия (ПД) Хочется надеяться, что мы с Вами разобрались в основных процессах, формирующих элементы «языка мозга» — нервные импульсы. Но нужно шагать дальше, ведь импульсы способны распространяться только по мембране одного нейрона. Для того чтобы закодированная их последовательностью информация могла быть передана следующему нейрону, требуется преодолеть щелевой контакт между возбудимыми клетками — синапс. А в синапсе принципы передачи и переработки информации будут не совсем такими, как на обычной электрогенной мембране (например, мембране аксона). Парадокс заключается в том, что мы не сможем понять принципы работы синапса, если не разберём ещё один важный момент в принципах работы аксональной мембраны — так называемые свойства потенциала действия. Принято выделять 5 базовых свойств ПД. 1. ПД подчиняется закону «всё или ничего» (рис. 33). Вообще говоря, приведённая схема так очевидна, что не требует дополнительных комментариев. Но сам по себе закон «всё или ничего» настолько интересен, что мы просто обязаны на нём остановиться и поговорить поподробнее. Ведь речь-то идёт об одном из базовых понятий и физиологии, и психологии: о порогах. Самое распространённое определение порога звучит весьма тривиально: порог — это то минимальное значение стимула, на которое живая система отвечает изменением своей активности. Принято различать абсолютные (верхний и нижний) и дифференциальные пороги (или пороги различения стимулов) (рис. 34). Рис. 33. Схема иллюстрирует закон «всё или ничего». Увеличивая силу наносимого от электростимулятора стимула, мы обнаруживаем, что при подпороговом значении стимула ПД не формируется, а развивается нераспространяющийся электротонический локальный ответ («ничего»). Если же мы перешагнули порог амплитуды стимула, разовьётся полноценный ПД, причём он будет одинаковым независимо от того, насколько нанесённый стимул сильнее порогового («всё»). Если говорить о ПД, то закон «всё или ничего» как раз характеризует особенности реагирования электрогенной мембраны нейрона на стимулы, превышающие или не превышающие уровень нижнего абсолютного порога. А вот дифференциальные пороги и их особенности нервного импульса как раз никак не касаются! Именно поэтому наивные попытки некоторых авторов вкладывать в форму, длительность, амплитуду и другие характеристики ПД какой-то информационный смысл свидетельствуют о том, что эти авторы никогда не видели импульсную активность нейронов «живьём» и знают о ней сугубо понаслышке. Давайте попробуем разобраться, почему же закон «всё или ничего» так императивен. Вспомните цикл Ходжкина. Для его запуска требуется преодолеть определённое пороговое значение деполяризации (в электрофизиологии его называют КУД — критический уровень деполяризации). Рис. 34. Понятие порогов чрезвычайно важно и для психологов, и для физиологов. Нижний абсолютный порог характеризует минимальное значение силы стимула, при котором живая система (клетка или её часть, ткань, орган, организм и т. д.) начинает «чувствовать» стимул (и его специфические особенности). Запуск ПД — это как раз игра на уровне нижнего абсолютного порога. Дифференциальные пороги характеризуют возможность различения силы (интенсивности) стимулов; чем больше сила стимула, тем превышения выше эти максимально пороги (закон возможной для Вебера — Фехнера). данной живой После системы интенсивности стимула начинает развиваться повреждение системы и утрачиваются индивидуальные признаки стимула; на сенсорном уровне это ощущается как боль. Этот уровень в некоторой степени индивидуален для каждого нейрона (разница иногда смешная — доли микровольта) и зависит исключительно от характеристик мембраны данной возбудимой клетки, а именно от количества (и, между прочим, качества) канальных белков. Давайте решим очень простую задачу: имеются два одинаковых по площади участка мембраны (1 квадратный микрометр, 1 квадратный метр — какая разница?), один весь буквально испещрён ионными каналами (пусть натриевыми), а на другом их ещё поискать придётся (рис. 35). Какую мембрану легче возбудить? Где нужен меньший начальный стимул? Где, стало быть, будет ниже КУД? Ответ-то простой: если на склоне горы лежат пять-шесть камней, потребуется сизифов труд, чтобы заставить их сымитировать хоть маленькую лавинку; если же весь склон в камнях — одного неосторожного движения (крика, слова) достаточно, чтобы сошла настоящая лавина (никогда так не делайте!). Рис. 35. Чем меньше плотность (количество на единицу площади) каналов на мембране, тем труднее эту мембрану возбудить: для мембраны Б потребуется значительно больший стимул, чем для мембраны А. Кстати, важным может оказаться не просто формальное количество каналов, а их способность активироваться: мы можем «отравить» мембрану каким-нибудь нейротропным препаратом (например, анестетиком типа ультракаина), и каналы-то на мембране останутся, а вот открыться в ответ на деполяризацию смогут далеко не все. Стало быть, способность мембраны ответить потенциалом действия на пришедший к ней стимул определяется тем, насколько она возбудима в настоящий момент (очень важно! мы скоро вернёмся к этому), и хватит ли силы стимула, чтобы запустить на ней цикл Ходжкина. Но, оказывается, судьба ПД зависит не только от силы, но и от длительности стимула. 2. Существуют критические для ПД параметры длительности стимула? Мы уже проводили аналогию между стимулом и стартёром автомобиля. Действительно, если аккумулятор сел, стартёру не хватает силы для запуска двигателя. Но если пытаться завести даже вполне исправную машину короткими поворотами ключа новоиспечённые водители), зажигания толку (как это тоже не будет: часто делают двигатель будет «глохнуть». И здесь тоже прямое сходство с ПД: существует не только пороговое значение силы стимула, но и пороговое значение его длительности (рис. 36). Причина та же: при слишком коротких стимулах не удаётся набрать «критическую массу» активированных каналов для запуска цикла Ходжкина. В реальной нервной системе, конечно же, отрабатывают оба эти параметра, и эту общую закономерность хорошо выражает так называемая «кривая силы — длительности» (рис. 37). Очень советую: постарайтесь с ней поработать — это хорошая возможность научиться читать графики. Рис. 36. Подпороговая длительность стимула так же, как и подпороговая сила, не провоцирует ПД, а вызывает только локальный ответ, который связан с тем, что в нашем теле все ткани являются хорошими проводниками электрического тока, а изоляторов, стало быть, просто нет. Поэтому при нанесении электрического стимула по мембране как простому физическому проводнику пойдёт ток, но «далеко не уйдёт», ведь вокруг-то тоже проводники (представьте себе оголённый провод, упавший в ванну с морской водой, — только экспериментировать не надо, а то весь район без света оставите!). Локальный — «locus=место»: где локальный ответ зародился, там (или почти там) и умрёт. Такие процессы называются электротоническими. В каждом ПД есть элемент локального ответа, но распространяются только потенциалы действия. Рис. 37. Классическая «кривая силы-длительности» графически выражает сразу оба свойства ПД. Обратите внимание на параллельные осям участки графика: параллельность оси X демонстрирует закон «всё или ничего» (стимул, меньший по силе порогового значения — реобазы, не вызовет ПД); параллельность участка графика оси Y показывает, что и для длительности стимула тоже существует пороговое значение, короче которого наносить стимул бесполезно (отсюда — «полезное время»). Эти показатели оказались не очень удобными, потому что характеризовали только уровни нижних абсолютных порогов. А ведь самой интересной является область перехода от одной параллельности к другой, когда и сила и длительность стимула закономерно дополняют друг друга. Для регистрации характеристик этой области было введено понятие хронаксии, которое отражает минимальную длительность стимула, вызывающего ПД при силе стимула, равной удвоенной реобазе. 3. Судьба ПД определяется также полярностью стимула (законы Пфлюгера) Ещё одним принципиально важным свойством ПД является его зависимость от полярности стимула. Вспомните наш электростимулятор (см. рис. 27): у него 3 главные ручки, позволяющие менять как раз силу, длительности и полярность стимула. То, что направление стимулирующего постоянного тока решительным образом влияет на возбудимость мембраны, впервые было показано ещё в позапрошлом веке замечательным немецким физиологом Пфлюгером в удивительно изящных для того времени экспериментах. Сформулированные Пфлюгером законы действия постоянного тока на возбудимые клетки и ткани звучат, на первый взгляд, довольно сложно даже в осовремененном вольном переводе: «При кратковременном действии постоянного тока на нервные клетки катод повышает возбудимость и может вызвать ПД, а анод снижает возбудимость и препятствует возникновению ПД». На самом деле, объяснение этих закономерностей достаточно примитивно и требует воспоминаний об уроках физики 5—6-го класса (рис. 38). Действительно, в этих эффектах постоянного тока очень мало биологического; практически обе закономерности проявляются на сугубо физическом уровне. И это очень хороший пример того, как физические (или химические) механизмы выстраивают логику сугубо биологических процессов. Не надо только увлекаться тем, что в философии называется редукционизмом: зачастую переход на более высокий уровень организации материи требует и перехода на иной уровень понимания процессов, обеспечивающих эту организацию, и сумма многих простых законов никак не складывается в один сложный закон (увы, физики и химики не всегда это понимают). Рис. 38. Законы Пфлюгера объясняются достаточно просто! В покое мембрана поляризована: внутри — минус, снаружи — плюс. Если мы внезапно, рывком (щёлкнули выключателем, нажали кнопку, etc) добавили к наружным плюсам катодные минусы, уровень поляризации резко уменьшится — то есть мембрана деполяризуется (дальше см. цикл Ходжкина). Если же к уже имеющимся наружным плюсам прибавить ещё и плюсы от анода, поляризация усилится, то есть возникнет гиперполяризация — состояние пониженной возбудимости мембраны хотя бы потому, что для запуска цикла Ходжкина придётся теперь преодолевать значительно больший энергетический порог, чем при нормальном уровне ПП. Интересно, что при выключении электростимулятора снятие анодного тока может (хотя и с меньшей вероятностью) привести к возникновению ПД. Механизм проверяется очень простым опытом: растяните пружину и отпустите. Если будет больно, сами виноваты — думать надо! Нужно коснуться ещё двух нюансов. Во-первых, если Вы внимательно читали предыдущие страницы, Вы, надеюсь, поняли, что для успешного запуска ПД на мембране требуется соблюдение трёх базовых условий: а) стимул должен быть надпороговой силы; б) стимул должен иметь надпороговую длительность; в) стимул должен быть катодическим. Эти сведения нам очень пригодятся для анализа механизмов синаптической передачи. Во-вторых, говоря о действии постоянного тока на мембрану нейрона, мы неслучайно подчёркивали кратковременность действия катода или анода. Дело в том, что при длительном действии тех же электрических полюсов на нейрон обнаруживаются Впервые это сумел иные, увидеть «непфлюгеровские» наш замечательный закономерности. соотечественник, представитель знаменитой в своё время уральской физиологической школы Бронислав Фортунатович Вериго. Вообще, вопреки распространённому (и упорно распространяемому в последние годы) мнению, российская физиологическая школа долгое время была (увы!) не менее уважаемой в научной среде, чем, например, химическая или физическая. Она дала миру такие великие имена, как Сеченов, Самойлов, Введенский, Вериго, Ухтомский, Павлов, Анохин, Бериташвили, Парин, Орбели, Черниговский, Костюк. Заглянув в учебник любой цивилизованной страны, Вы увидите там и «сеченовское торможение», и «доминанту Ухтомского», и «лёгочные рефлексы Парина», и «павловскую регуляцию пищеварения», и «парабиоз Введенского», и «катодическую депрессию Вериго», и «функциональные системы Анохина». Так вот, именно Вериго в начале ХХ века показал, что если катодический ток будет действовать на возбудимую клетку десятки миллисекунд и более, вместо возбуждения разовьётся глубокое торможение (депрессия), и способность генерировать ПД может быть утрачена. В то же время, он обнаружил и противоположный эффект — анодическую экзальтацию (правда, этот феномен довольно трудно воспроизводим и до сих пор вызывает дискуссии). Что же это за «вывернутые» закономерности? Оказывается, при долгом воздействии катода постепенно нарастает КУД. Мы-то с Вами теперь знаем, что критический уровень деполяризации зависит от свойств мембранных канальных белков, а вот первопроходцу, да ещё почти столетие назад, до этого надо было додуматься! Действительно, только сравнительно недавно, в последней трети ХХ века сложными современными методами удалось установить, что длительная деполяризация мембраны вызывает устойчивые нарушения проводящих свойств ионных каналов, что ведёт к нарастающему угнетению возбудимости. Да и вообще, возбудимость мембраны — это довольно лабильная характеристика. И рассмотрим мы это, в частности, на примере 4-го свойства ПД. 4. Потенциалы действия никогда не суммируются на мембране из-за наличия у неё свойства рефрактерности Придётся нам с Вами представить себе ещё один эксперимент. Для этого нашему электростимулятору придётся приделать ещё одну кнопку (и электронный блок соответственно), которая позволит подавать на нейрон не одиночные стимулы, а два подряд. Причём очень важно, чтобы мы могли постепенно увеличивать временной интервал между первым и вторым стимулами. Казалось бы, ну и что? В ответ на каждый стимул должен возникать свой ПД. А если интервал будет не 1—2 мс, а сотые или тысячные доли миллисекунды? Ни в одной, даже самой малограмотной (намёк на современные учебники Вы поняли) книжке Вы не увидите рисунков, на которых нервные импульсы слились бы в одно пятно. Всегда есть дискретность! Почему? Давайте смотреть (рис. 39). На рисунке — друг под другом два графика. Ось времени у них общая. Первый Вы, конечно же, узнали — это хорошо уже знакомый ПД. И хотя в этом издании большинство схем нарисовано с помощью простейших компьютерных приёмов, уверяем Вас — «живой» ПД на экране соответствующей регистрирующей аппаратуры будет выглядеть очень похоже. Рис. 39. Динамика возбудимости (способности генерировать ПД) мембраны нейрона (Б) при возбуждении (в процессе развития ПД) (А). Обратите внимание, что вслед за кратковременным небольшим повышением возбудимости (на начальном этапе деполяризации) наступает глубокий спад возбудимости (состояние рефрактерности), продолжающийся вплоть до завершения реполяризации. И только во время следовых процессов возбудимость возвращается к исходному уровню и колеблется около него. А вот второй график — это уже результат реконструкции «по экспериментальным точкам» целой серии последовательных опытов. В каждом опыте, напоминаем, мы должны чуть-чуть увеличивать интервал между первым и вторым стимулами. И получается удивительная вещь: обнаруживается относительно большой (это для нас 1 мс — незаметное мгновение, а для нейрона — целая вечность) отрезок времени, когда нейрон «не замечает» второй стимул, как будто его и не было. Этот период частичной или полной нечувствительности мембраны к наносимым стимулам получил название рефрактерности (относительной или абсолютной соответственно). На этой стадии развития ПД для того, чтобы мембрана отреагировала на второй стимул, он должен быть значительно сильнее, чем первый; а во время абсолютной рефрактерности стимулировать нейрон вообще бесполезно. Как Вы заметили, во время развития ПД есть два момента, когда возбудимость, наоборот, слегка повышена. Это начальный этап деполяризации (когда повторный стимул успевает «приплюсоваться» к первому — в самом деле, если мотор не заводится от стартёра, ему можно помочь заводной ручкой: вдвоём-то веселее!) и короткий момент следовой деполяризации (когда соотношение ионных градиентов ещё очень далеко от исходного). Вот мы и добрались, по существу, до базовой причины невозбудимости мембраны. Совершенно очевидно, что проблема в нарушении ионной проводимости. А в чём конкретно? Если Вы ещё не забыли, наряду с активационными воротами натриевые каналы имеют ещё и инактивационные. Если «залезть» на молекулярный уровень, белковые молекулы довольно инертны, и требуется время, чтобы они могли перейти из одного конформационного состояния в другое. Поэтому период, во время которого эти каналы не могут активироваться в ответ на новый стимул, достаточно значителен. Плюс к этому ещё и электрические характеристики мембраны во время деполяризации, овершута и реполяризации далеки от идеальных. В итоге реальная возбудимость мембраны в это время или крайне снижена (то есть пороги очень высоки), или отсутствует вообще. У этого свойства ПД есть два очень важных следствия. Первое: мы можем просчитать максимально возможные «скоростные» характеристики нейрона. Все, кому приходилось скачивать информацию с Интернета, хорошо понимают, насколько важной является «пропускная способность» канала связи. Наш мозг — это тоже информационная система (это ещё как сказать: «тоже»!), только в несчётное число раз более сложная и быстродействующая. Так как базовым мозговым кодом, по всей видимости, является пространственно-частотный, ясно, что максимально возможная для передачи по данному нейрону частота импульсов — чрезвычайно важный параметр. Для его обозначения наш великий соотечественник Введенский ввёл (извините за каламбур) 100 лет назад понятие лабильности. Лабильность — это максимальная частота импульсов, передаваемых или генерируемых нейроном в единицу времени. А теперь вернёмся к понятию рефрактерности (рис. 40). Если известно, что раньше завершения периода рефрактерности (то есть к началу следовых процессов) новый импульс возникнуть не может, мы можем рассчитать лабильность данного нейрона и единичный интервал принять за эталонный. Исходя из этого, определённый паттерн импульсной активности мы можем описать в двоичном коде, используя единицы на месте импульсов и нули там, где ПД теоретически мог бы быть, но не состоялся. Несколько десятилетий назад такие «двоичные игры» были весьма популярны среди нейробиологов, но потом стало ясно, что так просто задача «расшифровки мыслей» не решается. Сегодня очевидно, что это только очень малая толика истины и разгадки мозговых кодов надо искать в пространственновременных параметрах активности целых групп («ансамблей») нейронов или даже их отростков. Второе следствие из данного свойства ПД заключается в том, что при распространении импульсов вдоль мембраны они бегут поступательно от места возникновения и никогда не кружатся на одном месте. Здесь тоже «виновата» рефрактерность, но к анализу этого следствия мы перейдём в рамках следующего, пятого свойства потенциала действия. Рис. 40. На схеме А показано, что повторный ПД может возникнуть только «на выходе» из реполяризации первого ПД (точка 2а) или позже (точки 2б и 2в), но никак не раньше, то есть суммация ПД на мембране одного нейрона невозможна (вследствие рефрактерности). Схема Б имитирует возможный механизм двоичного кодирования в таком нейроне (1 — есть импульс, 0 — нет импульса, но мог бы быть в соответствии с лабильностью). 5. ПД распространяются по мембране бездекрементно (то есть без угасания) Действительно, если мы вспомним механизм цикла Ходжкина, то сразу увидим способность нервных импульсов распространяться по мембране, ведь в каждое следующее мгновение открываются всё новые и новые натриевые каналы. А где их взять? Да на соседнем же участке мембраны, естественно. И причин для затухания ПД нет, пока не кончится электрогенная мембрана. Казалось бы, что тут обсуждать? Оказывается, очень даже есть что! Дело в том, что в нашей нервной системе присутствуют два типа проводящих импульсы волокон (напоминаем, что волокна — не более, чем отростки нервных клеток: аксоны или дендриты). Более древние — немиелинизированные (или безмякотные), и эволюционно более молодые — миелинизированные. И принципы бездекрементного распространения ПД по ним существенно различаются. В немиелинизированном нервном волокне ПД распространяются с помощью механизма, очень похожего на движение волны по воде (если бы ещё эту волну в каждое мгновение кто-то подгонял, ведь энергия растрачивается). Представьте себе три последовательных этапа этого процесса (рис. 41). Рис. 41. Три «стоп-кадра», которые демонстрируют последовательные этапы распространения нервного импульса по немиелинизированному волокну. К сожалению, на бумаге очень трудно передать движение, динамику. Поэтому напрягите Ваше воображение, чтобы увидеть, как вспыхивающий в каждой новой точке мембраны ПД за счёт местных (локальных) токов деполяризует соседние участки, запуская там цикл Ходжкина. Этим токам «всё равно», что деполяризовать: ту мембрану, которая находится впереди, на пути распространяющегося ПД, или ту, по которой импульс только что прокатился. Но ведь есть 4-е свойство ПД, и благодаря рефрактерности остающийся «за спиной» участок мембраны не отвечает на деполяризацию. Поэтому ПД и распространяется вдоль нервного волокна, а не кружится на одном месте. В первый момент времени на невозбуждённый участок мембраны поступает нервный импульс. За счёт местных токов он деполяризует соседний участок, запуская на нём цикл Ходжкина. Через активировавшиеся ионные каналы возникает лавинообразно нарастающий натриевый ток, что приводит к увеличению деполяризации мембраны вплоть до реверсии потенциала. Этот «вновь рождённый» ПД деполяризует следующий участок мембраны и т.д. Нервный импульс поступательно движется по мембране аксона от аксонного холмика до пресинаптического окончания, не угасая. Прямо скажем, это очень надёжный механизм проведения сигнала, но, увы, довольно медленный: скорость проведения нервных импульсов по таким волокнам колеблется в пределах 0,3 — 15 м/с (в зависимости от толщины волокна, как Вы помните). Казалось бы, не так уж и медленно? Много ли в нашем теле метровых волокон? И тем не менее, для выживания в постоянно меняющихся условиях такой скорости, видимо, недостаточно. Такой забавный пример: если бы идущий от сетчатки глаза вглубь мозга зрительный нерв состоял из немиелинизированных волокон, то для получения той же скорости проведения информации, которая имеется у нас в реальности, потребовался бы нервный ствол диаметром… около 50 см! Представьте свои глазки на таких стебельках (точнее, пеньках) и поймёте, что что-то здесь не так. Из чего же «сделан» настоящий человеческий зрительный нерв (кстати, он по толщине вполне сопоставим с просветом стержня от Вашей шариковой ручки)? В том-то и дело, что он состоит из быстропроводящих миелинизированных нервных волокон, скорость проведения импульсов в которых может достигать 120 м/с. Существенная разница, не правда ли? За счёт чего же возможен такой выигрыш? Попробуем разобраться. Для этого вспомним строение миелинизированного нервного волокна (рис. 42). В отличие от безмякотного в нём мембрана через определённые промежутки покрыта муфтами из жироподобного вещества — миелина, спирально намотанного на поверхность волокна (которое, кстати, в этом случае называется осевым цилиндром). Рис. 42. По миелинизированному нервному волокну ПД распространяются сальтоторно, то есть «прыжками» от перехвата к перехвату Ранвье. Так как осевой цилиндр покрыт миелиновыми муфтами, под которыми трансмембранные ионные токи маловероятны, ПД «перескакивает» от перехвата к перехвату сугубо электротонически (вспомните локальный ответ!) — практически мгновенно, но с очевидным затуханием. Если бы не сверхвысокая возбудимость мембраны в перехватах Ранвье (а плотность ионных каналов там на два порядка выше, чем в обычной мембране), судьба нервных импульсов в таких волокнах была бы плачевна. Но эволюция ухитрилась найти такие параметры частоты перехватов и возбудимости, что миелинизированные волокна проводят ПД весьма надёжно и с высокой скоростью: ведь потери времени на ионные токи здесь остались только в перехватах. Изображённые пунктиром ПД должны проиллюстрировать идею электротонического затухающего проведения — это скорее образ, чем результат регистрации. Эту работу по созданию миелиновых муфт выполняют сопутствующие нейронам шванновские клетки. Миелин — вовсе не электрический изолятор, как жизнерадостно пишут в последние годы авторы некоторых учебников. Повторю, что, к их огорчению, в нашем живом теле нет истинных изоляторов, так же как и надёжных проводников. Функция у этих муфт иная: на покрытых ими участках мембраны до крайности затруднён трансмембранный перенос ионов и, стало быть, нет механизма, способного обеспечить запуск цикла Ходжкина. В результате мы видим гениальную находку природы: в миелинизированных волокнах нервные импульсы распространяются между перехватами Ранвье (оголёнными участками мембраны, находящимися между муфтами миелина) скачками, сальтоторно (от латинского глагола saltore — прыгать, скакать, кувыркаться). По существу, это электрический пробой. Учитывая миниатюрность нейронов, можно считать, что на эти скачки время вообще не тратится. Однако здесь есть небольшая неприятность: при электротоническом проведении потенциалы неуклонно гаснут по мере удаления от источника. Но эта проблема устраняется за счёт чрезвычайно высокой возбудимости мембраны в перехватах Ранвье. В них плотность (то есть количество на единицу площади) ионных каналов очень высока (в 100 раз больше, чем в обычной мембране!), и даже почти совсем угасшего потенциала достаточно, чтобы запустить на такой мембране цикл Ходжкина. В результате время при сальтоторном проведении ПД тратится только на перекачку ионов через каналы в перехватах, и скорость распространения нервных импульсов в миелинизированных волокнах может быть на порядок (даже больше) выше, чем в немиелинизированных. Маленькое заключение. Мы с Вами очень кратко рассмотрели пять базовых свойств ПД. Хочется ещё раз напомнить, что обратились мы к ним прежде всего потому, что эти знания лягут в основу понимания процессов биоэлектрогенеза в самых главных — с информационных позиций — узловых точках нейронной сети: синапсах. К этому вопросу мы и перейдем в следующей лекции. 4.4. Постсинаптические потенциалы Надеюсь, в общих чертах строение синапса Вам знакомо (рис. 43). Рис. 43. Реконструкция синапса, на которой представлены его основные компоненты. Сейчас же хотелось бы вкратце напомнить, что под синапсом понимают щелевой контакт между нейроном и другими возбудимыми клетками (не только нервной, но и мышечной или эндокринной), в котором происходит передача и переработка (!!!) информации (рис. 44). Пока мы не будем касаться особенностей работы электрических синапсов. Если бы мы были разумными ракообразными, это была бы наиважнейшая тема, но для нас, млекопитающих, гораздо важнее синапсы химические, потому что именно они формируют сложнейший информационный аппарат нашего мозга. Рис. 44. Среди нескольких вариантов межнейрональных синапсов (аксоаксональ-ные, аксо-дендритные, дендро-аксональные и т. д.) мы для анализа выбрали аксо-соматический синапс. Причём на этой нашей схеме магистральный синапс (по горизонтали) «отягощён» ещё двумя подобными ему. Будем считать, что сотни остальных остались где-то за плоскостью картинки. Обратите внимание на важную деталь: характеры импульсации в левом (передающем) нейроне и в правом (принимающем) существенно различны. То, что было недопустимым на уровне нервного волокна (изменение импульсного сигнала), становится чуть ли не обязательным для синаптической передачи. Напоминаю, что на каждом из 10 11 — 1012 нейронов, составляющих нашу нервную систему, в среднем сходится около 1000 синапсов. Каждый синапс — весьма сложно организованный аппарат, в котором имеется масса процессов и механизмов, большинство из которых просто бесполезно для простой передачи сигнала. Вся история научного поиска учит, что природа до чрезвычайности ленива и никогда не будет создавать сложную систему там, где можно было бы обойтись простой. Это просто невыгодно, и эволюция с лёгкостью вымывает, элиминирует все «архитектурные излишества». Стало быть, очень усложнённая структура синапса призвана обслуживать и очень сложную функцию, а это никак не передача сигнала (вот уж для чего электрические синапсы оптимальны!), а именно его обработка. Вот давайте и попробуем для начала проанализировать, какие особенности электрических процессов в химическом синапсе позволяют нейронам перерабатывать, а не просто пересылать информацию. Как работает химический синапс? Для начала вспомним, как он устроен (рис. 45). Рис. 45. Крайне упрощённая схема синапса, в которой из реального межнейронного контакта остались только принципиально значимые для нашего обсуждения элементы. Если на первых этапах не вдаваться в особо тонкие детали, то главными элементами химического синапса являются: - пресинаптическое окончание (пресинапс), на электрогенной мембране которого имеются потенциалзависимые кальциевые каналы (там есть и другие каналы, но пока в эти подробности вдаваться не будем), а внутри пресинапса расположены мембранные везикулы (пузырьки) с молекулами медиаторов. Непосредственно к обращённой в синаптическую щель пресинаптической мембране примыкают сократительные белки (SNAREбелки). отметим, что в каждом пресинапсе может присутствовать до 10000 везикул, в каждой из которых может быть до 10000 молекул разных (это принципиально важно!) медиаторов; - синаптическая щель, разделяющая передающий и принимающий информацию нейроны. Ширина этой щели около 40 нм (для сравнения: толщина клеточной мембраны около 20 нм); - постсинаптическая мембрана, на которой расположены рецепторные белки, способные с высоким сродством связываться с медиаторами и другими специфическими лигандами (например, ядами или лекарствами), и управляемые этими рецепторами лигандзависимые ионные каналы. Безусловно, эта схема сугубо примитивно характеризует чрезвычайно сложный в реальности синапс, но для описания базового алгоритма его работы она достаточна. А теперь сам короткий алгоритм (буквально по пунктам) передачи информации от нейрона к нейрону: По электрогенной мембране (например, аксона) ПД достигает пресинапса (тем, кто уже забыл, как распространяются нервные импульсы по электрогенной мембране, неплохо бы отлистать эту книжку на несколько страниц назад). Так как механизм распространения ПД включает в себя деполяризацию мембраны, то и мембрана пресинапса тоже деполяризуется. В результате этой деполяризации активируются потенциалзависимые кальциевые каналы. В этом смысле они очень напоминают уже знакомые нам потенциалзависимые каналы, пропускающие по концентрационному градиенту ионы натрия и калия при генерации ПД. Ионы кальция по градиенту концентрации проникают в пресинапс. Вообще о кальции стоит поговорить подробнее: это чрезвычайно «популярный» в биологических системах катион. Он абсолютно необходим и для прочности костей (без кальция их хоть узлом завязывай!), и для мышечных сокращений, и для системы свёртывания крови, и для иммунной системы, и для работы сердца, и т. д. Вот и синапс без ионов кальция работать не будет. Кальций вызывает активацию сократительных белков пресинапса, и они дружно, синхронно притягивают везикулы с медиаторами к пресинаптической мембране, обращённой в синаптическую щель. И снова есть необходимость в уточнениях. Дело в том, что сократительные белки пресинапса (так называемые SNARE-белки) и по химическим свойствам, и по механизмам функционирования чрезвычайно похожи на сократительные белки нашей поперечно-полосатой мускулатуры. Поэтому имеет смысл чуть более детально остановиться на том, как работают наши скелетные мышцы (рис. 46). Главными рабочими элементами каждого мышечного волокна являются фибриллярные (нитчатые) белки: актин и миозин. Миозин значительно крупнее и тяжелее, чем актин. Эти белки расположены в мышечном волокне квазипараллельно, подобно зубцам двух расчёсок, направленных навстречу друг другу. Уникальной особенностью актина и миозина является их способность динамически взаимодействовать образования временных химических «мостиков». друг с другом путём А В Рис. 46. Схематическое изображение процесса сокращения мышечного волокна. А — локальный участок (фрагмент саркомера) мышечного волокна в покое: между квазипараллельными фибриллярными молекулами сократительных белков актина и миозина расположен блокирующий белок тропомиозин, препятствующий взаимодействию этих белков. В — деполяризующие мембрану мышечного волокна (в том числе и мембрану поперечных трубочек) потенциалы действия опосредованно активируют (пока ещё не совсем ясно, как именно, хотя гипотезы есть вполне красивые) кальциевые каналы терминальных цистерн продольных трубочек, и ионы Ca2+ по градиенту концентрации выходят из продольных трубочек в область сократительных белков; в результате тропонин (белок-сателлит тропомиозина) связывается с ионами Ca2+, снимая тропомиозиновый блок; между освобождёнными нитями актина и миозина возникает динамическое взаимодействие: формируются временные мостики, перемещающиеся вдоль нитей (подобно рукам участников двух команд, перетягивающих канат) и обеспечивающие скольжение нитей актина и миозина навстречу друг другу. Если внимательно приглядеться, можно увидеть, что на схеме В Z-пластинки расположены ближе друг к другу, чем на рис. А, то есть саркомер укоротился (мышца сократилась). Кстати, наши мышцы могут укорачиваться на 50% своей длины. Эти мостики пробрасываются от миозина к актину, быстро меняют угол своего прикрепления (белки «подтягиваются» за них навстречу друг другу), отсоединяются, перемещаются вперёд, снова подтягиваются и т. д. В результате нити сократительных белков начинают скользить друг навстречу другу, и мышца сокращается. Естественно, что этот механизм «скользящих нитей» весьма энергозатратен, — ведь мы хорошо знакомы с усталостью после физической работы. В детали этих энергетических процессов мы вдаваться не будем — они выходят за рамки данного раздела. Гораздо важнее для нас, почему же это происходит не всегда, а только «по приказу» от мотонейрона? Дело в том, что в покое нити актина и миозина разобщены блокирующим белком тропомиозином и их взаимодействие невозможно. Для снятия этого блока нужно, чтобы тропомиозин изменил свою конформацию, а это, в свою очередь, достигается при взаимодействии ещё одного белка — тропонина (белка-саттелита тропомиозина) с ионами Ca2+. Тропонин — типичный представитель довольно обширного семейства кальцийсвязывающих белков (ещё один яркий пример такого белка — кальмодулин, играющий роль второго посредника, то есть внутриклеточного переносчика информации. Наберитесь терпения — об этом поговорим чуть позже). И вот отсюда становится понятной роль кальция: высвободившись в результате деполяризующего действия ПД из терминальных цистерн продольных трубочек (вполне тривиальные для клетки мембранные структуры — вспомним комплекс Гольджи, эндоплазматический ретикулум и т. п.), он диффундирует к сократительным белкам, связывается с тропонином и таким образом снимает блокировку с акто-миозинового взаимодействия. Значит, активация сократительных белков происходит не случайно, не спонтанно, а исключительно под влиянием ПД, то есть управляемо. А теперь вернёмся к пресинапсу. Аналогия между SNARE — белками и сократительными белками мышечного волокна — пусть и не абсолютно точная, но вполне допустимая. И кальций здесь отрабатывает ту же роль. Только задача в пресинапсе другая: не обеспечить сокращение нейрона (ещё не хватало, чтобы «шевеление мозгами» происходило буквально!), а добиться одновременного, сугубо синхронизированного притягивания множества везикул с медиаторами к пресинаптической мембране. То есть опять же — процесс высвобождения медиаторов идёт не спонтанно, не хаотично (такое, впрочем, случается, в своё время мы поговорим о так называемых МПКП), а управляемо, в ответ на приход ПД в пресинапс. При взаимодействии пресинаптической мембраны с везикулярными мембранами везикулы лопаются, и молекулы медиаторов высвобождаются в синаптическую щель (экзоцитоз). Этот процесс тоже заслуживает особого комментария (рис. 47). Рис. 47. Схема экзоцитоза. Везикулы, заполненные молекулами медиаторов, сталкиваются с пресинаптической мембраной и вскрываются в синаптическую щель. В результате множество (около 10000 в каждой везикуле) молекул медиаторов оказывается в узком (до 40 нм) щелевом пространстве между передающей и принимающей информацию клетками. В принципе, слияние двух липидных мембран при их простом соприкосновении — процесс, термодинамически вполне естественный. Представьте себе две янтарные капельки жира в тарелке с горячим (!!) бульоном. Достаточно согнать их ложкой вместе, слегка ткнуть этой же ложкой вдоль получившейся восьмёрки, и на Ваших глазах из двух капель получится одна большая. Отработает гидрофобность жирнокислотных «хвостов» липидных молекул, категорически не желающих общаться с водной фазой. Однако наш мозг — всё же не бульон с капельками жира, поэтому процесс экзоцитоза посложнее, чем только что описанный. Сегодня открыта целая команда мембранных белков, принимающих в нём участие, да и ионы кальция нужны для активации не только SNARE-белков, но и самого экзоцитоза. К сожалению, эти интересные детали далеко выходят за рамки нашего курса. Высвободившиеся из везикул молекулы медиаторов диффундируют через синаптическую щель к постсинаптической мембране. Нужно заметить, что за один цикл (то есть в ответ на один ПД) из пресинапса одновременно высвободится содержимое примерно 10000 везикул, то есть около 100000000 молекул медиаторов. Молекулы медиаторов с высоким сродством («высокоаффинно») связываются со своими специфическими рецепторами и активируют их. И этот процесс заслуживает особого рассмотрения (рис. 48). Нам ещё предстоит серьёзный разговор о многочисленных медиаторах и других информонах. Однако сейчас имеет смысл приглядеться к самым важным проблемам лиганд-рецепторных взаимодействий. И прежде всего это принцип комплементарности, согласно которому медиаторы связываются сугубо со «своими» рецепторами (например, ацетилхолин холинорецептором, адреналин — с адренорецептором, и т. д.). — с Рис. 48. Принцип комплементарного связывания медиаторов сугубо со «своими» рецепторами на первый взгляд прост и примитивен. «Ключ к замку», «рука к перчатке» — всем знакомы такие метафоры, описывающие этот принцип (например, антиген-антитело, фермент-субстрат и т. п.). Нужно всего лишь, чтобы две взаимодействующие молекулы зеркально, подобно пазлам, подходили друг к другу (пространственно, энергетически, ещё какнибудь…). И, казалось бы, при современных возможностях физикохимических методов ничего не стоит доказать эту идею. Однако мы не имеем пока прямых доказательств и опираемся только или на математические модели, или на косвенные данные (например, результаты фармакологического анализа). Есть ещё и проблема чисто теоретического плана: в последние десятилетия открыто множество биологически активных веществ (в первую очередь, медиаторов), способных высокоспецифично связываться с белковыми сайтами мембраны, то есть рецепторами. Прогресс в этой области явно не собирается тормозиться, так что уже скоро счёт таких веществ (рецепторных лигандов) пойдёт на сотни, а то и на тысячи. Возникает резонное недоумение: сколько же генетической памяти надо потратить, чтобы закодировать этакую прорву специализированных рецепторных белков?! Нет у нас такого запаса ДНК — на все случаи жизни! Так что исследовать ещё и исследовать… Механизмы таких лиганд-рецепторных взаимодействий достаточно неплохо изучены, но их детализация выходит за рамки этого издания. Второй важный момент заключается в том, что активация рецептора (то есть изменение его энергетики и конформации) происходит мгновенно, именно в тот миг, когда идет прямое химическое взаимодействие крохотной молекулы медиатора и громадного рецепторного белка. Активированные рецепторные белки, в свою очередь, активируют систему передаточных (G-) белков, которые одним из двух возможных способов (рис. 49, 50) открывают лигандзависимые ионные каналы постсинаптической мембраны. Первый способ получил название «быстрого», при нём рецептор и канал — это, по сути, единый комплекс, и активация ионотропных рецепторов (способных только активировать ионные каналы) приводит к быстрому и непродолжительному открыванию ионных каналов постсинаптической мембраны. Второй способ — «медленный»: активированные медиаторами метаботропные рецепторы (способные влиять на обмен веществ — метаболизм — в клетке) запускают систему II мессенжеров, которые, в свою очередь, стимулируют цепочку биохимических реакций, приводящих в конечном итоге к фосфорилированию мембранных канальных белков; в результате ионные каналы открываются медленнее, но открытыми могут оставаться существенно дольше, чем в первом случае (секунду, минуты и даже часы). Есть необходимость сказать два слова о II мессенжерах. Рис. 49. На чрезвычайно упрощённой схеме представлены два принципиальных механизма активации лигандзависимых ионных каналов. А — «быстрый» способ, который реализуется при активации медиатором ионотропных рецепторов постсинаптической мембраны. Так как данные рецепторы являются, по существу, элементами ионного канала (напомню, что каналы — это надмолекулярные белковые комплексы, состоящие из нескольких субъединиц белка), процесс активации происходит практически мгновенно, без потерь времени. Б — «медленный» механизм, при котором активация рецептора запускает систему II мессенжеров (это внутриклеточные передатчики информации; речь о них пойдёт чуть позже), которые способны активировать различные метаболические каскады внутри клетки, в том числе процесс фосфорилирования (обозначено на схеме буквой Р) канальных белков. В результате лигандзависимые каналы переходят в активированное состояние и могут оставаться открытыми не несколько миллисекунд (как в «быстром» способе), а секунды, минуты и даже часы. Рецепторы, запускающие такой механизм, получили название метаботропных. Ещё раз подчеркну, что схема не учитывает очень многие детали процесса активации каналов (например, участие в нём G-белков и фосфолипидов мембраны) и иллюстрирует только главные этапы. Когда в начале 60-х годов прошлого века Сазерленд открыл первый из вторых мессенжеров (именно «вторых», а не «вторичных», как по недомыслию закрепилось в нашей литературе) — циклический аденозинмонофосфат (цАМФ), это стало настоящим событием! К этому времени давно уже были открыты межклеточные химические коммуникаторы: гормоны (первый в истории гормон — секретин — был открыт Бейлиссом и Старлингом в далёком 1902 году) и медиаторы (здесь приоритет принадлежит Отто Леви, который в 1920 году доказал медиаторную функцию ацетилхолина, за что был удостоен Нобелевской премии). Рис. 50. Более совершенная в художественном смысле реконструкция ионотропных и метаботропных рецепторов и управляемых ими каналов, показанных на рис. 49. Открытие Сазерленда стало следующим шагом в понимании химических механизмов передачи информации в живых системах. За без малого полвека удалось обнаружить не так уж много II мессенжеров. Кроме цАМФ, это его «собрат» — ещё один циклический нуклеотид цГМФ (циклический гуанозинмонофосфат), кальцийсвязывающий белок кальмодулин, некоторые фосфолипиды мембраны (например, инозитол-3-фосфат, диацилглицерол) и, что стало научной сенсацией конца ХХ века, совсем уж простые молекулы типа оксида азота (NO). Несмотря на невеликое количество этих внутриклеточных посредников, их роль в работе клеток чрезвычайно значительна. В следующем издании я надеюсь познакомить Вас с этим более подробно, но уже сейчас возьмите на заметку, что чашка чаю или кофе бодрит именно потому, что кофеин стимулирует систему II мессенжеров. Однако продолжим путешествие по синапсу. Что за каналы открываются «по приказу» рецепторов? Это очень важно понять. так как на постсинаптической мембране ионные каналы принципиально отличаются от тех, которые мы рассматривали при анализе ПД, характер ионных токов, и соответственно потенциалов, будет очень существенно отличаться от тех, которые формируют нервные импульсы: а) во-первых, каналы постсинаптической мембраны являются лигандзависимыми, а не потенциалзависимыми, то есть эти каналы активируются исключительно «по команде» рецептора и принципиально неадекватно реагируют на изменения потенциала, поэтому цикл Ходжкина на них запустить невозможно; б) во-вторых, эти каналы значительно менее селективны, чем потенциалзависимые, поэтому через них возможны даже встречные потоки ионов. Например, на постсинаптической мембране различают Na +—K+каналы и Cl-— K+-каналы (рис. 51, 52); в) в-третьих, длительность активированного состояния таких каналов значительно больше, чем у потенциалзависимых (для каналов, активированных ионотропными рецепторами, больше примерно в 5 раз, а для управляемых с метаботропных рецепторов — секунды, минуты и даже часы!). Рис. 51. На постсинаптической мембране главную роль играют два типа лигандзависимых каналов: Na+—K+ и Cl–—K+. Они значительно менее селективны, чем потенциалзависимые (причём определяющими для изменения потенциала мембраны являются потоки ионов натрия и хлора, а не калия), и гораздо дольше остаются в открытом состоянии. Исходя из этих особенностей каналов постсинаптической мембраны, сами постсинаптические потенциалы (ПСП) также вопиюще не совпадают по своим свойствам с потенциалами действия (табл. 1): 1) ПСП не подчиняются закону «всё или ничего», так как на постсинаптической мембране нет субстрата для его воспроизведения — потенциалзависимых каналов. Поэтому ПСП относятся к электротоническим процессам и способны меняться градуально: чем больше ионных каналов активировалось, тем больше (по модулю) будет сам потенциал. Рис. 52. Очень, на мой взгляд, выразительные схемы двух видов (и двух подвидов) ионных каналов, управляемых физическими (А) и химическими (В) стимулами. А — потенциалзависимые (слева) и механозависимые (справа) каналы; В — слева показан канал, активирующийся медиатором, справа — активирующийся через запуск системы II мессенжеров. 2) длительность ПСП всегда значительно больше, чем у ПД (минимум в 5 раз). 3) в отличие от ПД, ПСП могут быть как деполяризационными (в случае активации Na+—K+-каналов), так и гиперполяризационными (если активируются Cl–—K+-каналы). Таким образом, во-первых, мы видим, что уже по первым трём свойствам градуальные постсинаптические потенциалы принципиально отличаются от потенциалов действия. Таблица 1. Сравнительная характеристика потенциалов действия (ПД) и постсинаптических потенциалов (ПСП) Потенциалы Постсинаптические действия потенциалы Возникают вследствие активации Возникают при активации потенциалзависимых ионных лигандзависимых ионных каналов каналов Подчиняются закону «всё или Не подчиняются этому ничего»: возникают только при закону и могут меняться надпороговых значениях стимула и градуально (в зависимости всегда одинаковы от соотношения ионных токов) Чрезвычайно кратковременны Значительно (более, чем в 5 (1—2 мс) раз) длительнее Однополярны: всегда начальная ВПСП — деполяризация деполяризация с последующей (соответствует катоду); ТПСП гиперполяризацией — гиперполяризация (соответствует аноду) Никогда не суммируются на Есть пространственная и мембране одного нейрона временная суммация как на внутрисинаптическом, так и межсинаптическом уровне Распространяются бездекрементно Электротонические за счёт перманентного потенциалы: угасают в связи «самовозрождения» через цикл с отсутствием изоляторов в Ходжкина живых организмах Во-вторых, самое время вспомнить, что при рассмотрении первых трёх свойств ПД мы обсуждали прежде всего условия его генерирования (напомню: стартовый стимул должен быть надпороговым по амплитуде и по длительности, а также катодическим). С учетом «задач» ПСП (обеспечить возникновение ПД на электрогенной мембране) особенности его свойств становятся всё более актуальными — не для имитации ли постсинаптических процессов существуют электростимуляторы, а? Подумайте об этом! Однако вернёмся пока к нашему алгоритму. 4) Как Вы, надеюсь, помните, потенциалы действия не суммируются на одном волокне (из-за наличия рефрактерности). Для ПСП таких запретов не существует: убедительно доказано, что постсинаптические потенциалы могут суммироваться, причём эта суммация может быть как пространственной, так и временной. Следовательно, к градуальности каждого ПСП добавляется ещё и возможность суммации множества потенциалов, возникших более-менее одновременно на нескольких соседних синапсах. Причём такая суммация может привести и к наращиванию ПСП (если суммируются ВПСП), так и к его вырождению (когда в процесс вмешиваются ещё и ТПСП). 5) Наконец, в отличие от ПД, постсинаптические потенциалы распространяются с угасанием, ведь на постсинаптической мембране нет субстрата для формирования цикла Ходжкина (и соответственно бездекрементного проведения) — потенциалзависимых ионных каналов. А вот теперь самое главное: мы можем попытаться понять, каким образом возникающие на постсинаптической мембране Na+, K+ и Cl– ионные токи формируют постсинаптические потенциалы, и как затем эти ВПСП/ТПСП, многократно суммируясь и неизбежно угасая, обеспечивают возникновение разного количества ПД на электрогенной мембране за пределами синапсов (прежде всего в аксональном холмике нейрона). По существу, можно повторить наше обсуждение механизмов генерации ПД, в ходе которого мы подавали начальный стимул с помощью электростимулятора. Напомню, что этот стимул должен быть надпороговой амплитуды (силы), надпороговой длительности и обязательно катодическим. Теперь представьте себе, что в результате суммации ВПСП и ТПСП у нас сформировался некий ПСП, отвечающий всем этим требованиям, то есть сумма ВПСП оказалась заведомо больше суммы ТПСП. Можем ли мы быть уверены в том, что теперь за возникновение ПД на электрогенной мембране принимающего нейрона можно не беспокоиться? Увы, нет. Потому что электротонический характер ПСП не даёт никаких гарантий по поводу того, будет ли достигший электрогенной субсинаптической мембраны суммарный постсинаптический потенциал надпороговым для генерации там ПД или угаснет до подпорогового уровня. 11. Казалось бы, вопрос о синаптической передаче информации исчерпан. Исходный набор перекодирований, нервных импульсов, преобразовался в пройдя конечный через набор, несколько как правило, отличающийся от исходного. И механизмы этих перекодирований болееменее понятны. Но есть ещё одна немаловажная деталь в алгоритме работы синапса. В 7-м пункте нашего обсуждения мы говорили о том, что медиаторы, высокоспецифично связываясь с мембранными рецепторами, активируют их. Сам процесс активации с энергетических позиций выглядит достаточно очевидным: конформация белковой молекулы (а рецептор, напомню, — белок) в большой мере зависит от её энергии, то есть при прочих равных условиях белок «выбирает» такую(-ие) конформацию (-ии), в которой его молекула обладает минимумом свободной энергии. При взаимодействии маленькой молекулы медиатора с огромной рецепторной молекулой энергетическое состояние белка-рецептора на короткий миг меняется, и соответственно меняется и его конформация. Этого мига достаточно, чтобы активировать дальнейшие процессы в мембране (вспомним G-белок!) и передать информацию по цепочке дальше. А что же медиатор? Ведь он свою роль уже исполнил? Что ж, «мавр сделал своё дело и мавр может уйти». Даже не может, а должен! Должен, потому что импульсы поступают в пресинапс один за другим, выделяются всё новые и новые порции медиаторов, а с чем эти молекулы будут связываться, если рецептор ещё занят? Чтобы такого не случилось, существует несколько молекулярных «дворников», которые должны быстро очистить рецептор от отработавшего медиатора. Трёх главных «блюстителей чистоты» Вы сейчас увидите. 1) Ферментативное расщепление медиатора (рис. 53). Взаимодействие медиатора с рецептором вызывает активацию фермента, как правило, связанного с постсинаптической мембраной. Фермент катализирует реакцию распада молекулы медиатора, и рецептор очищается. Так, например, происходит инактивация самого известного из медиаторов — ацетилхолина. Рис. 53. Благодаря ферментативному катализу молекула медиатора распадается на отдельные компоненты непосредственно в синаптической щели. Компоненты утилизируются (в том числе могут пойти на синтез новых молекул медиаторов), а очищенный рецептор готов к приёму следующей порции медиатора. Именно с нарушением этого процесса связаны психические нарушения, вызываемые такими ингибиторами фермента ацетилхолинэстеразы, как дихлофос, хлорофос, пресловутый «novichok» и другие препараты для борьбы с вредными членистоногими (тараканами, клопами и др.). 2) Обратный захват медиатора (рис. 54). После взаимодействия медиатора с рецептором возвращают запускается медиатор в система белков-транспортеров, пресинаптическое окончание которые (своего рода «безотходное производство»). Рис. 54. Чрезвычайно упрощенная иллюстрация механизма обратного захвата медиаторов (например, моноаминов) или их отдельных фрагментов. Трудно представить, что, управляя этим процессом, можно вытащить человека из тяжёлой депрессии. Однако это факт… 3) Интернализация рецептора. Этот сравнительно недавно открытый механизм очистки рецептора от отработавшего медиатора может лечь в основу объяснения одного из базовых механизмов наркотической толерантности/зависимости. Поэтому рассмотрим его более детально (рис. 55). Интернализация рецептора представляет собой очень быстрый (по некоторым данным, несколько микросекунд) процесс, реализующийся через несколько этапов: медиатор связывается с рецептором и активирует его; рецептор транслирует сигнал в клетку, а медиатор становится для него «обузой», от которой надо избавиться; рецептор вместе с медиатором и частью липидного бислоя «ныряет» в цитоплазму и замыкается в микровезикулу; Рис. 55. Упрощенная схема процесса интернализации рецептора. Показаны последовательные этапы: активированный медиатором рецептор, успев передать сигнал дальше, «тонет» под мембрану; в подмембранном пространстве медиатор подвергается ферментативному расщеплению, а освободившийся от него рецептор возвращается на мембрану. Нарушение этого процесса (на этапе ферментативного расщепления) при наркомании является одной из базовых причин толерантности/зависимости. на комплекс «медиатор-рецептор» набрасываются ферменты (учитывая, что интернализация характерна для рецепторов к медиаторам пептидной природы, это пептидазы) и расщепляют молекулу медиатора; освободившийся от медиатора рецептор «всплывает» на мембрану (сразу же или в ответ на новый синаптический сигнал) — он готов к новым информационным подвигам! Маленькое заключение. Таким образом, мы вкратце рассмотрели базовый механизм передачи сигнала в химическом синапсе (рис. 56). Рис. 56. Вы видите последовательность событий в чрезвычайно примитивном химическом синапсе, в котором присутствует только один вид медиатора. Активация постсинаптической мембраны происходит только через ионотропные рецепторы, а натриевый ток через каналы постсинаптической мембраны формирует исключительно ВПСП. В реальных синапсах всё, как правило, гораздо сложнее… По существу, мы убедились, что закодированная последовательностью нервных импульсов информация, как минимум, три раза меняет в процессе синаптической передачи свой материальный «субстрат»: а) первое преобразование происходит в пресинапсе, когда ПД провоцирует выделение медиаторов, и информация, образно выражаясь, переносится на химический носитель — медиатор; б) второе перекодирование постсинаптических — потенциалов это формирование вследствие активации градуальных медиаторами рецепторов и последующей активации ионных каналов постсинаптической мембраны; в) и третий шаг — генерация новой последовательности нервных импульсов в аксонном холмике под влиянием постсинаптических потенциалов всех активированных на текущий момент синапсов. Вместо заключения Надеюсь, прочитанные Вами страницы хоть в некоторой степени позволили Вам представить себе, какие электрические процессы в нейронах обеспечивают передачу и переработку информации в нервной системе. Если так — значит, книжка писалась не зря! Напоминаю, что это пособие написано для тех, кто делает первые шаги к пониманию работы мозга (не так важно, кто это: физики, психологи, математики, биологи…). Поэтому и нарочито упрощённый язык, и ограниченный круг вопросов, которые здесь рассмотрены. Ведь даже просто история изучения «живого электричества», начавшаяся несколько веков назад с Л. Гальвани и А. Вольта, — это уже бесконечно увлекательная сага о великих находках, искренних заблуждениях и цистернах пролитого пота (и крови, между прочим, правда, в основном крови экспериментальных животных). Сегодняшний день электробиологии — это гимн точности измерений на самых разных уровнях, и прежде всего на микроуровне (очень модная сегодня приставка «нано-» — это ведь как раз про клеточную и молекулярную нейробиологию: здесь и нанометры, и нановольты, и наномоли). А там, где измерено, там в конечном итоге и понято! Однако рискну напомнить, что мозг — это не только электрическая, но и химическая «машина». Поэтому не советую расслабляться: следующий раздел — про медиаторы, гормоны и другие информоны — уже ждет вас. Надеюсь, прочитанные вами страницы хоть в некоторой степени позволили вам представить себе, какие электрические процессы в нейронах обеспечивают передачу и переработку информации в нервной системе. Если так — значит, эту главу вы читали не зря! Напоминаю, что учебное пособие написано, в основном, для тех, кто делает первые шаги к пониманию работы мозга (не так важно, кто это: психологи, физики, математики, упрощённый язык, и биологи…). ограниченный Поэтому круг вопросов, и нарочито которые здесь рассмотрены. Ведь даже просто история изучения «живого электричества», начавшаяся несколько веков назад с Л. Гальвани и А. Вольта, — это уже бесконечно увлекательная сага о великих находках, искренних заблуждениях и цистернах пролитого пота (и крови, между прочим, правда, в основном крови экспериментальных животных). Сегодняшний день электробиологии — это гимн точности измерений на самых разных уровнях, и прежде всего на микроуровне (очень модная сегодня приставка «нано-» — это ведь как раз про клеточную и молекулярную нейробиологию: здесь и нанометры, и нановольты, и наномоли). А там, где измерено, там в конечном итоге и понято! Однако рискну напомнить, что мозг — это не только электрическая, но и химическая «машина». Поэтому не советую расслабляться: следующая глава — про медиаторы, гормоны и другие информоны — уже на следующих страницах. Ну и самое сейчас главное. По сути, мы с вами уже можем назвать, как минимум, 7 мишеней для действия фармакологических агентов: - ионные каналы, - ионные насосы, - синтез медиаторов, - высвобождение медиаторов, - рецепция медиаторов, - функционирование II мессенжеров, и - очистка рецепторов от медиаторов. Как видите, как минимум в четырех случаях из семи мишенями являются главные информоны нервной системы – медиаторы. Именно о них (и не только) пойдет речь в следующей главе, из которой вы увидите, что наш мозг – не только «электрическая машина», но и «химическая машина». ИНФОРМОНЫ: МЕДИАТОРЫ, ГОРМОНЫ, II МЕССЕНЖЕРЫ В предыдущей главе мы разобрали основные принципы передачи информации в организме человека и животных с помощью электрических сигналов. Таким образом, мы выяснили, что мозг (да и весь организм в целом) является своеобразной «электрической машиной». Но если следовать машинной аналогии (подарок аж из XVIII века), то в такой же – если не в большей – степени мы являемся «химической машиной». Дело даже не в том, что все без исключения функции (и структуры, кстати) нашего организма есть результат химических реакций. Нас же, напомню, интересует относительно узкий вопрос: передача информации и возможность вмешательства в эту передачу. Естественно, без химической «кухни» здесь не обойтись. Вспомните: все электрические потенциалы в нервной системе, с которыми вы теперь знакомы, являются порождением сугубо химических процессов. А уж про синаптическую передачу и говорить нечего – это, в основном, сугубо химический процесс (точнее, цепочка химических реакций). Если к этому добавить внутриклеточные химические процессы, работу всей эндокринной системы, которая неразрывно связана с нервной, химическую коммуникацию между отдельными животными внутри вида (вспомните феромоны, аттрактанты и т.п.) или на межвидовом уровне (алломоны), становится понятно, что без рассмотрения многообразия химических взаимодействий наши знания о механизмах передачи информации в живых системах будут явно ущербными. И снова хочу повторить, что в рамках этой книжки мы не сможем погружаться в тонкости этих механизмов. Эта задача – для других изданий (очень советую, например, обратиться к фундаментальному пособию «Нейрохимия», вышедшему под редакцией академика И.П. Ашмарина). Наша скромная цель – увидеть основные принципы функционирования химических механизмов передачи и обработки информации. К ней и будем стремиться. Начнём с основных понятий. Первым из них является введённое в мировую практику уже упоминавшимся здесь генералом Игорем Петровичем Ашмариным понятие информонов. На мой взгляд, трудно найти более точное и при этом короткое название для той массы веществ, которые обеспечивают передачу информации в живых системах. Под этот термин подходят и внутриклеточные переносчики информации, и межклеточные (здесь далеко не только медиаторы), и те вещества, которые передают информацию между особями (табл. 2). Таблица 2. Некоторые классы информонов (по: [Ашмарин, 1996], с изменениями). Класс информонов II мессенжеры Уровень передачи информации Внутриклеточный Медиаторы Межклеточный Гормоны Гистогормоны Межтканевой Межклеточный Феромоны Алломоны Нуклеиновые кислоты Внутривидовой Межвидовой Системный Типичные примеры цАМФ, цГМФ, кальмодулин, NO, c-fos Ацетилхолин, норадреналин, ГАМК, энкефалин, глутамат Инсулин, тироксин, кортизол Простагландины, лейкотриены, тромбаксаны Половые аттрактанты самок Зоотоксины ДНК, иРНК, тРНК, АТФ, ГТФ Большинство классов мы разберем в этой главе. Очевидно, что начать надо с медиаторов. Классические медиаторы Прежде всего, почему их называют «классическими». Это тот случай, когда история науки вступила в противоречие с историей жизни на Земле. Сегодня сложилось устойчивое мнение, что в эволюции первыми информонами были пептиды, причем их начальная функция у примитивных одноклеточных заключалась в передаче информации на внутри- и межвидовом уровне. В дальнейшем, с формированием многоклеточных организмов, эти пептиды стали выполнять функции гормонов и медиаторов. Лишь существенно позднее, у высших многоклеточных, к классу медиаторов присоединились химически более простые молекулы, по масштабу не превышающие одиночную аминокислоту. Однако в истории нейронаук именно они стали первыми из открытых медиаторов и, вследствие этого, получили название классических. И первым был открыт ацетилхолин. Ацетилхолин, холинорецепторы, ацетилхолинэстераза Медиаторные свойства ацетилхолина были открыты в 1921 году австрийским физиологом Отто Лёви в результате тонких (для того времени) экспериментов на сердце лягушки. Однако само вещество было обнаружено на 7 лет раньше будущим Нобелевским лауреатом и будущим сэром Генри Дейлом (закономерно, что Нобелевскую премию Лёви и Дейл получили вместе – Рис. 58. Отто Лёви и структурная формула ацетилхолина. разобраться в 1936 году), когда он пытался в странных физиологических эффектах спорыньи. Надо сказать, что Лёви проявил завидное научное упрямство, занявшись доказательством медиаторных функций именно ацетилхолина. В начале ХХ века доминировало утверждение, высказанное другом Дейла - Томасом Элиоттом: первым открытым медиатором будет адреналин. И большинство крупнейших нейрохимиков того времени с безнадежным упорством пытались это подтвердить. Пройдет не одно десятилетие, пока это, действительно, не будет доказано, и тогда станет очевидным, что существовавшие в 10-20-х годах ХХ века методы и подходы просто не могли привести к желаемому результату. Во-первых, центральными (то есть действующими внутри мозга) медиаторами окажутся ближайшие химические родственники адреналина норадреналин и дофамин, а адреналин больше представлен в периферических синапсах. Во-вторых, время жизни адреналина в синапсе составляет всего несколько миллионных долей секунды, что не могло позволить «выловить» его с помощью методов, использовавшихся сто лет назад. Но вернемся к Отто Лёви. История его открытия – находка для психоаналитика. Надо сказать, что долгие месяцы его эксперименты тоже преследовали сплошные неудачи, и он был близок к отчаянию (я опираюсь на его воспоминания). И вдруг однажды ему приснился сон, в котором он увидел всю необходимую схему эксперимента. Проснувшись среди ночи, он схватил карандаш и клочок бумаги, записал все увиденное и, совершенно счастливый, улегся досыпать. Наутро выяснилось, что он не может разобрать ни строчки. Весь день он безуспешно пытался в лаборатории воспроизвести обрывки сна, и отправился спать в абсолютно разбитом состоянии. И ему снова приснился тот же самый сон!! Теперь он уже не стал ждать милостей от природы, накинул халат и в домашних тапочках устремился в лабораторию (благо, бежать было не так уж и далеко – в соседнее здание). Приснившийся эксперимент был воспроизведен и, как позже писал сам Лёви, «в четыре утра я понял, что я – Нобелевский лауреат». Не правда ли, отличный пример влияния доминирующей мотивации на наше сознание и подсознание! Пресловутый Зигмунд Фройд – отдыхает. Я имею ввиду, естественно, мотивацию, направленную на решение задачи, а вовсе не на получение премии… Суть поставленного Лёви решающего эксперимента следующая. От двух лягушек (усыпленных, не расстраивайтесь) он взял два сердца. Одно из них он поместил в физиологический раствор (простейший физиологический раствор – 0,9% раствор обычной поваренной соли) и стал стимулировать блуждающий нерв. Электростимулятор, надеюсь, вы еще не забыли. Блуждающий нерв или вагус (n.vagus)– это главный нерв парасимпатической нервной системы, активация которого замедляет сердцебиение. После этого он взял этот физиологический раствор и добавил его в емкость, где находилось сердце второй лягушки. И оно тоже стало биться медленнее! То есть впервые удалось доказать, что при стимуляции блуждающего нерва выделяется вещество, заставляющее сердце биться реже. Немного позже выяснили, что этим веществом является именно ацетилхолин – первый медиатор. По сути, в итоге именно Отто Лёви открыл эру медиаторов, которая продолжается до сих пор. Естественно, за прошедшее столетие об ацетилхолине стало известно почти все. Оказалось, что это главный медиатор нервно-мышечных синапсов. Мы о таких синапсах немного уже говорили. Именно через них «приказ» о сокращении мышечных волокон доводится до них от мотонейронов спинного мозга (или ствола головного мозга, если речь идет, например, о мышцах, обеспечивающих моргание). Отсюда получается, что любое наше движение прямо зависит от ацетилхолина. И не только движение: наши дыхательные мышцы тоже являются поперечнополосатыми, стало быть, возможность дышать прямо связана с активностью ацетилхолина. Об этом много веков назад «узнали» индейцы нынешней Гвианы (это на Амазонке) и стали использовать для охоты яд местной лианы Strychnos toxifera, который потом, уже после открытия Америки, получил название «яд кураре». Думаю, любители Конан-Дойля, Агаты Стрихнос ядоносный Кристи и Честертона узнали это вещество, столь часто используемое разнообразными злодеями. Достаточно маленькой капельке этого яда попасть в кровь, и тапир, пума, человек, крокодил или анаконда перестанет двигаться, а потом и дышать. А причина выяснилась только после открытия Лёви: яд кураре блокирует рецепторы к ацетилхолину (холинорецепторы) в нервно-мышечных синапсах. Но во всяком открытии можно найти и хорошее, и плохое. Сегодня во всем мире серьезные операции хирурги проводят с использованием производного от яда кураре – лекарства dтубокурарина. Это позволяет существенно уменьшать травмирование тканей при хирургическом вмешательстве и, соответственно, облегчать выздоровление. Но управление движениями – далеко не единственная задача ацетилхолина. Ацетилхолин является, по существу, главным медиатором вегетативной нервной системы, управляющей – напоминаю – нашими внутренними органами. Так как печальный опыт учит, что эта важнейшая управляющая система организма часто оказывается где-то на периферии внимания, вынужден напомнить, в чем ее главные особенности. Вегетативная нервная система, в отличие от соматической, занимается управлением внутренними органами. Иногда ее называют автономной, потому что нормальному человеку (про йогов пока не говорим) не позволено сознательно регулировать работу, например, сердца. Напомню, что вегетативная система делится на симпатическую и парасимпатическую. Сразу хочу развеять распространенное заблуждение. Так как типичным примером активности симпатической и парасимпатической систем является работа сердца, многие наивно обобщают, что парасимпатическая система всегда тормозит работу всех внутренних органов, а симпатическая – активирует. Это совсем не так. Например, работа нашего пищеварительного тракта, наоборот, активируется вагусом (напомню – главным парасимпатическим нервом - блуждающим), а тормозится симпатическими нервами. Тогда, может быть, их действие просто противоположно? – И тоже не совсем точно. Ближе всего к истине другое объяснение. Парасимпатическая система обеспечивает, в основном, тоническую регуляцию (поддерживает тонус), а симпатическая – фазическую, то есть быстрые ответные реакции на требования окружающей среды. Не случайно симпатическая система является одним из главных компонентов физиологической системы стресса. Теперь снова посмотрите на рисунок. Обратите внимание, что в вегетативной системе, в отличие от соматической, сигнал от мозга к мышцам доставляется не одним нейроном (помните – мотонейрон?), а двумя последовательными. Первый несет информацию от мозга к ганглиям, то есть «нервным узлам», и называется преганглионарным (пре- - перед). Второй нейрон начинается в ганглии, и заканчивается во внутреннем органе, гладкими мышцами которого он управляет. Этот нейрон называется постганглионарным (пост- - после). Есть редкие исключения, но на них мы останавливаться не будем. Этот механизм объединяет симпатическую и парасимпатическую системы. А теперь о различиях. Прежде всего, центральные нейроны симпатической системы расположены в спинном мозгу, а парасимпатической – в стволе головного мозга, и в самых нижних (крестцовых) отделах спинного мозга. Ганглии тоже расположены по-разному. Симпатические – или в околопозвоночных нервных цепочках (их называют паравертебральными: пара- - около, вертебра – позвоночник), или отдельно, немного дальше от спинного мозга (их называют превертебральными). Вам, конечно, знаком самый большой превертебральный ганглий – это солнечное сплетение. Ганглии парасимпатической системы «ближе к народу»: они расположены или рядом с иннервируемым органом (называются экстрамуральными: экстра- - снаружи), или в его стенке (интрамуральные: интра- - внутри). И еще одно важное различие, непосредственно касающееся темы медиаторов. Ацетилхолин является главным медиатором преганглионаров и парасимпатической, и симпатической системы. То есть во всех ганглиях синаптическая передача информации осуществляется преимущественно именно ацетилхолином. Кроме того, ацетилхолин обеспечивает передачу информации с постганглионаров на управляемый орган, но только в парасимпатической системе. В симпатических постганглионарах главными медиаторами являются адреналин и норадреналин, но о них речь у нас впереди. А здесь – самое основное: в трех из четырех ключевых узлов передачи информации в вегетативной нервной системе главным медиатором является именно ацетилхолин. Кроме того, ацетилхолин представлен и в головном мозгу: прежде всего, в стволе головного мозга, гипоталамусе и коре больших полушарий. Ацетилхолин участвует в регуляции и переключении основных функциональных состояний: сон – бодрствование, быстрый сон (точнее фаза быстрого движения глаз) – медленный сон (опять же, точнее – медленноволновая фаза, потому что во время ее развития регистрируются медленные изменения потенциалов нейронных ансамблей, так называемые дельта-волны ЭЭГ). Да и уровень бодрствования у нас нестабилен. Наверное, вы замечали, что труднее всего заниматься примерно с 2 до 5 после полудня. Это потому, что человек – единственное дневное животное, отказавшееся от дневного сна. Но ацетилхолин об этом «не догадывается», и пытается уговорить нас немного подремать. Вообще ацетилхолин, как мы уже обсуждали, осуществляет фазическую регуляцию. Например, при стрессе он стремится понизить уровень возбуждения, а при апатии – наоборот, повысить. Еще один очень важный вопрос связан с мембранными рецепторами к ацетилхолину – холинорецепторами. В середине прошлого века выяснилось, что у этих рецепторов постсинаптической мембраны, кроме ацетилхолина, могут быть и другие агонисты (напомню: агонисты стимулируют рецепторы и заставляют их открывать лигандзависимые ионные каналы, в отличие от антагонистов, которые эти рецепторы блокируют). Оказалось, что для одних холинорецепторов агонистом является никотин, а для других – мускарин. Про никотин – понятно, это та самая беда всех курильщиков. А вот мускарин – это алкалоид всем известного гриба мухомора. Первые назвали nхолинорецепторами (никотинчувствительными), вторые – m- холинорецепторами (мускаринчувствительными). Довольно скоро выяснилось, что эти рецепторы сильно отличаются друг от друга. Если nхолинорецепторы оказались ионотропными (вспоминайте, что это значит, - мы это проходили), то m-холинорецепторы – метаботропные, то есть работают через систему мессенжеров. вторых Соответственно, эффекты первых проще, а вторых существенно сложнее. Это проявляется и в вегетативной нервной системе, и в центральных эффектах. Например, активация n-холинорецепторов снижает выраженность стресса, успокаивает, способствует концентрации внимания. Не подумайте только, что я пропагандирую табакокурение! Полезность этих эффектов никотина полностью перекрывается тем разрушительным действие, которое он оказывает, прежде всего, на сосудистую систему. Облитерирующий эндартериит – это страшное заболевание кровеносных сосудов, прежде всего, сосудов ног. Вызывается оно, прежде всего, никотином, который постепенно разрушает регуляцию сосудистого тонуса. На последней стадии «лечение» хирургическое: ногу отрезают как колбасу – все выше и выше. Пока нога не кончится… Мускаринчувствительные рецепторы тоже представлены и в периферической, и в центральной нервной системе. Они участвуют в реализации процессов памяти, научения, обслуживании базовых инстинктов. Не случайно всяческие берсерки и викинги использовали «мухоморовку» как допинг перед битвой. Глупо, но понятно: все равно помирать, так пусть уж с музыкой. Даже незначительная передозировка мускарина ведет к нарушениям сознания, галлюцинациям и, в конечном итоге, к смерти. Каждый год любители исторических реконструкций времен викингов и, конкретно, Одина (бог такой был у них) пополняют городские кладбища. И наконец, вопрос об очистке холинорецепторов от отработавшего медиатора. Она происходит по первому механизму из трех, разобранных нами. Фермент ацетилхолинэстераза (или проще – холинэстераза) гидролизует, то есть расщепляет ацетилхолин, освобождая от него холинорецептор. Механизм простой и довольно распространенный. Особенность этого вопроса в том, что существует целая группа веществ, ингибирующих (то есть подавляющих) этот фермент. Прежде всего, это фосфорорганические соединения (ФОС). Поэтому призываю вас: не увлекайтесь всяческими дихлофосами и хлорофосами! На их упаковках не зря написано, что это яд. То, что букашки-таракашки умирают от них быстрее, чем мы, объясняется просто. Всякий яд действует по принципу: миллиграмм яда на килограмм веса жертвы. Таракан, клоп или колорадский жук несколько легче нас, поэтому помрет первым. Но и только! Я вполне понимаю те трудности, которые могли возникнуть у вас при чтении этой подглавки. Но на примере первого в истории науки медиатора я хотел показать вам хотя бы малую толику тех многочисленных сложностей, с которыми сталкиваются нейрохимики и нейрофармакологи. Следующие разделы, уверяю вас, будут несколько проще. Биогенные амины: адреналин, норадреналин, дофамин, серотонин и гистамин адреналин норадреналин дофамин серотонин гистамин Прежде всего, эту группу классических медиаторов объединяет тот факт, что все они являются производными аминокислот. Не удивляйтесь: аминокислоты – это не только «бусинки», из которых строятся короткие пептиды и длинные белки. У аминокислот много других обязанностей в организме, и передача информации в синапсе – одна из них (об этом – в следующей подглавке). А кроме того, они могут быть «сырьем» для синтеза медиаторов. Так как в аминокислотах всегда есть аминогруппа (NH2), и их производные тоже имеют одну группу (это моноамины: адреналин, норадреналин, дофамин и серотонин) или даже две группы (диамин – гистамин). Начнем которые с тех моноаминов, объединяются в род катехоламинов. Это дофамин, норадреналин и адреналин. Именно в таком порядке, потому что все они являются производными широко распространенной аминокислоты тирозина. Как видно из схемы, каждое преобразование идет в одно «химическое действие», каждом этапе и на реакции катализируют разные ферменты. Не удивляйтесь, что на завершающем этапе вы видите загадочные «норэпинерин» и «эпинефрин». Это же все те норадреналин и адреналин, только в переводе не с латинского, а с греческоого англичане отличие языка. и от предпочитают именно Почему-то американцы, остального в мира, использовать греческие по происхождению термины. А еще обратите внимание на промежуточный продукт этой цепочки реакций: из тирозина образуется ДОФА (дигидроксифенилаланин), а уже из него – дофамин. Это нам еще пригодится. Дофамин является классическим центральным медиатором, то есть передает информацию в синапсах мозга. Причем представительство вырабатывающих дофамин нейронов в мозгу очень ограничено. Их общее количество – всего около 400 тысяч (сравните с десятками миллиардов!). Главные локусы - это аркуатное (дугообразное) ядро гипоталамуса, вентральная покрышка и черная субстанция среднего мозга. Особый интерес представляют нейроны черной субстанции (s. nigra), аксоны которых тянутся к такой структуре базальных ганглиев, как полосатое тело (corpus striatum), включающее хвостатое и чечевицеобразное ядра. Эти нейроны образуют так называемую нигростриатную систему. Нейроны гипоталамических ядер, в свою очередь, направляют свои аксоны в структуры лимбической системы, в фронтальную и префронтальную кору и в гипофиз. Естественно, существуют специализированный мембранные белки, связывающиеся с дофамином – Dрецепторы. Функции дофамина во многом могут быть выведены методом «от противного»: какие функции организма нарушаются при недостатке или избытке дофамина. Одним из распространенных психоневрологических расстройств является синдром Паркинсона (паркинсонизм). Во всем мире паркинсонизмом сегодня страдает около 4 миллионов человек – это второе место после болезни Альцгеймера. Болезнь проявляется тремором (дрожанием) конечностей, скованностью и замедленностью движений, неспособностью к тонкой моторике, нарушениями памяти и психическими расстройствами. Главной причиной паркинсонизма считается снижение содержания дофамина в нигростриатной системе, связанное с гибелью дофаминсодержащих нейронов черной субстанции. Поэтому одной из первых попыток лечения этой болезни была терапия дофамином. Однако быстро выяснилось, что дофамин плохо проникает через гемато- энцефалический барьер (ГЭБ - барьер между кровью и мозгом, не пропускающий в мозг большинство присутствующих в крови молекул и защищающий его от всевозможных нейротоксинов) и, самое главное, быстро превращается в норадреналин и адреналин. Так что больной не только не перестает дрожать, но еще и начинает это воспринимать крайне агрессивно. И тогда фармакологов посетило озарение! А что, если в схеме синтеза дофамина отступить на шаг назад? То есть вводить не дофамин, а его предшественника – ДОФА. В результате в аптеках можно купить эффективное лекарство – леводопу (L-DOPA). Нет, уже погибшие нейроны, конечно, не восстанавливаются, но с помощью препарата можно стабилизировать состояние больного. С другой стороны, нарушения нормального распределения дофамина по структурам мозга может быть причиной шизофрении. Показано, что повышение уровня дофамина в полосатом теле при понижении его уровня в префронтальной коре часто наблюдается при разных формах шизофрении. Поэтому в психиатрической клинике довольно популярной является терапия с помощью галоперидола – блокатора (антагониста) дофаминовых рецепторов. Вдумайтесь, отчего могут зависеть нарушения уровня дофамина в мозгу? – Конечно, нельзя исключить генетическую предрасположенность, но главное объяснение лежит в повреждениях ферментных систем: если будет плохо работать тирозингидроксилаза (вернитесь к схеме синтеза катехоламинов), дофамина будет не хватать, если же сбой произойдет в функционировании ДОФА-декарбоксилазы, мы получим патологический избыток дофамина, но, соответственно, дефицит норадреналина и адреналина. Но вернемся к доказательствам «от противного». Теперь, зная, какие неприятности несет недостаток или избыток дофамина, мы сможем коротко сформулировать – каковы его главные функции. Дофамин участвует в регуляции движений: и тонких, точных, и стереотипных – то есть сложных двигательных навыков. Дофамин играет важную роль в таких когнитивных и аффективных функциях, как мотивации, эмоции, научение (особенно – эмоциональное), память. Эмоциональная составляющая здесь далеко не случайна, ведь мозговым центром эмоций и мотиваций является лимбическая система, на которую воздействуют дофаминсодержащие нейроны гипоталамуса. Был в истории науки период, когда дофамин называли «гормоном счастья». Оказалось, неправда! Выяснилось, что это медиатор, а не гормон, и к счастью он никакого отношения не имеет. А заблуждение было связано с тем, что дофамин был обнаружен в структурах так называемой «системы вознаграждения» (reward system) мозга, которую вначале окрестили «центром удовольствия». Только никакой это не центр удовольствия, а сложная нейронная сеть, которая отвечает за мотивацию поискового поведение и активизацию процесса оценки, изучения новых, неожиданных факторов и свойств среды. Теперь давайте перейдем к адреналину и норадреналину. Их принято называть «гормонами стресса». Во многом это правда, но не во всем. Да, их главная роль проявляется, действительно, при стрессе. Да, и адреналин, и норадреналин выделяются из мозгового слоя надпочечников, действуя на клетки-мишени после транспортировки кровью, как и положено гормонам. Но оба эти вещества одновременно являются и медиаторами. Вокруг медиаторных свойств адреналина до сих пор ведутся споры, но большинство данных указывает на его участие в синаптической передаче в гипоталамусе и в симпатических постганглионарах, прежде всего, иннервирующих сердце и сосуды. Под его влиянием активируется выработка некоторых рилизинггормонов гипоталамуса (чуть позже мы о них поговорим), усиливается работа сердца (частота сокращений и их мощность), повышается артериальное давление. Норадреналин также представлен в симпатических постганглионарах, но он является и центральным медиатором, в частности, в ретикулярной формации ствола головного мозга. Здесь он активирует так называемую систему бодрствования, которая позволяет нам оставаться в тонусе, несмотря на усталость. Он является главным медиатором голубого пятна (locus coeruleus) среднего мозга. Аксоны нейронов этого ядра распространяются вниз - в спинной и промежуточный мозг, - и вверх – вплоть до коры больших полушарий. На мембранах клеток имеются разные рецепторы к этим катехоламинам, главными из которых являются так называемые α- и β-адренорецепторы. Они в разных соотношениях присутствуют во всех тканях нашего тела, благодаря чему суммарное действие адреналина и норадреналина затрагивает все системы организма. Вместе с гормонами мозгового слоя надпочечников эти катехоламины как медиаторы симпатических постганглионаров формируют симпатоадреналовую систему, являющуюся одной из главных в реализации стресса. Обратите внимание, как всего два довольно простых вещества обеспечивают целый комплекс реакций в организме, направленных но один главный результат: усиление работы мозга и мышц и обеспечение этого необходимой энергией. Напомню: гликолиз, гликогенолиз и липолиз – это расщепление глюкозы, гликогена и липидов («жиров») в присутствии кислорода для «зарядки» клеточных аккумуляторов – АТФ. Все остальные звенья представленной ниже схемы направлены на быструю доставку энергоресурсов до мозга и мышц, ведь на первой стадии стресса (стадии тревоги) нам нужно или бежать, или драться (есть, правда, третий вариант – замереть, - но об этом позже). Сегодня принято следующее определение стресса: стресс – это неспецифическая, защитная, стадийная, системная, редуцированная психофизиологическая реакция на повреждение или его угрозу. И в рамках этой концепции симпатоадреналовая система обеспечивает первую – самую энергозатратную – стадию стресса: стадию тревоги. Безусловно, это не единственные функции адреналина и норадреналина. Они работают и в относительно спокойных, не стрессогенных контекстах, сопровождая двигательные, когнитивные и эмоциональные нагрузки. Серотонин (его другое название – 5-гидрокситриптамин, в англоязычных текстах – 5-НТ) также является производным аминокислоты – триптофана. Первоначально он был выделен из слизистой оболочки кишечника, затем – из сыворотки крови. Поэтому решили, что это гормон. А так как повышение уровня серотонина было зафиксировано при эйфории, а понижение – при депрессии, его тут же объявили «гормоном счастья» (еще одним!). И снова довольно скоро выяснилось, что это не только гормон, но и медиатор, да и со счастьем тоже как-то не складывается. Как центральный медиатор, серотонин, в основном, локально представлен в стволе головного мозга и в гипоталамусе. На периферии его представительство гораздо шире – и в кишечнике, и в матке, и в системе крови и кровообращения, и т.д. Сегодня обнаружено уже 7 типов 5-НТ-рецепторов (да еще несколько подтипов), которые обеспечивают многообразие функций серотонина. Среди них есть и ионотропные, и метаботропные, и возбуждающие, и тормозные. Если сосредоточить внимание на психических проявлениях деятельности серотонина, можно увидеть, что его недостаток приводит к депрессии, а избыток – к галлюцинациям. Не случайно само открытие серотонина было связано с исследованием функций ДЛК (диэтиламида лизергиновой кислоты, по-немецки – LSD; использование в русской транскрипции аббревиатуры ЛСД столь же неприлично, как обозначение бывшего лидера мировой экономики сокращением УСА!) и других галлюциногенов: например, мескалина, буфотенина и псилоцибина. Наконец, переходим к краткому обсуждению диамина – гистамина. Гистамин тоже образуется из аминокислоты – гистидина. В мозгу гистамин вырабатывается небольшими группами нейронов мамиллярных тел гипоталамуса. Предполагается, что именно гистамин является главным медиатором, который связывает нервную систему с иммунной. На периферии он представлен и в легких, и в пищеварительном тракте, и в печени, и в покровных тканях (коже, слизистых). Вместе со своими Н-рецепторами (их известно 4 типа) вырабатывающие гистамин клетки образуют так называемый тучноклеточный аппарат, отвечающий за развитие воспаления – защитной реакции, которая развивается у нас в ответ на различные повреждения: ранения, инфекции. Не случайно тучные клетки расположены, в основном, в тех тканях, которые обращены во внешнюю среду. Видимо, именно тесной связью гистамина с системой иммунитета можно объяснить его важную роль в аллергических реакциях. Поэтому самыми распространенными противоаллергическими препаратами являются антигистаминные средства, то есть антагонисты Н-рецепторов. Индивидуальные аминокислоты: глутамат, аспартат глицин аспартат, глицин, ГАМК глутамат ГАМК Аминокислоты являются не только строительными элементами для белков и пептидов или предшественниками медиаторов предыдущей группы, но выполняют в организме множество других функций. В частности, некоторые из них сами могут выступать в роли медиаторов. Аспартат (аспарагиновая кислота; окончание «–ат» обозначает кислоту) и глутамат медиаторами, (глутаминовая то есть их кислота) являются возбуждающими рецепторы активируют Na+-K+-каналы постсинаптической мембраны. Глутамат и аспартат присутствуют в качестве сопутствующих медиаторов в подавляющем большинстве синапсов мозга. Соответствующие рецепторы – AMPA-, NMDA- и каинатные – являются самыми распространенным в нейронных сетях центральной нервной системы. Эта особенность несколько десятилетий назад послужила причиной изящной, но не подтвердившейся гипотезы «стартовых медиаторов». Предполагалось, что эти медиаторы (прежде всего – глутамат) первыми связываются со своими рецепторами при активации синапса и таким образом «предупреждают» постсинаптическую мембрану о скором прибытии других медиаторов, то есть готовят постсинаптическую мембрану к предстоящей работе. На самом деле объяснение, судя по всему, нужно искать в другой области. В том, что нейроны являются самыми «прожорливыми» клетками. Мозг, как известно, потребляет около 30% всей энергии, вырабатывающейся в нашем организме. Причем питание (все ту же глюкозу и кислород) нейронам доставляют окружающие их глиальные клетки, и «кормят» они нейроны только после того как нейроны сработают, то есть передадут информацию. Именно поэтому известная байка о 7% (5%, 10% и т.д.) нейронов, которые мы используем в течение жизни, не выдерживает критики. Это что же, 93% (95%, 90% и т.п.) нейронов у нас гибнет? – Чушь! Как же глиальные клетки «понимают», что этот нейрон поработал и заслужил свой обед? – Оказалось что на глиальных клетках тоже есть мембранные рецепторы, и эти рецепторы, в основном, к глутамату. То есть, когда синапс (и, соответственно, нейрон) сработал, часть глутамата «вытекает» за пределы синаптической щели и сигнализирует глиальным клеткам о нейронной активности. Так что в мозгу «кто не работает, тот не ест». Вторая пара медиаторных аминокислот – глицин и ГАМК (γаминомасляная кислота) – имеет рецепторы, которые активируют Cl--K+каналы постсинаптической мембраны, что, как вы, надеюсь, помните, вызывает формирование не ВПСП, а ТПСП. То есть обе эти аминокислоты – тормозные медиаторы. Это принципиально важно, ведь еще полтора века назад наш великий земляк Иван Михайлович Сеченов показал, что в нервной системе всегда присутствует не только возбуждение, но и торможение. И обнаружение тормозных медиаторов явилось прекрасным этому подтверждением. Глициновые рецепторы, как и рецепторы к ГАМК (ГАМКА, ГАМКВ или ГАМКС) очень широко распространены в мозгу, что доказывает участие этих аминокислот в большинстве нервных процессов. Собственно говоря, на этом мы можем закончить знакомство с классическими медиаторами и перейти к несравнимо более обширному классу нейропептидов, которые почти всегда являются и медиаторами, и гормонами. Нейропептиды Прежде всего, давайте вспомним, что такое пептиды. Если опускаться до совсем уж бытового уровня, можно сказать, что это очень короткие белки. Пептиды тоже представляют собой «нитку бус», в которой каждая из «бусинок» - аминокислота (точнее – аминокислотный остаток). Принципиальное отличие заключается в том, что белки сворачиваются в сложные клубки, образуя третичную структуру (вспомните школьное – «конформация белка»). Пептиды слишком коротки, чтобы сворачиваться, и, как правило, представляют собой линейные молекулы. Синтез пептидов идет по тем же принципам, что и синтез белков. Схемы строения всех наших белков и пептидов хранятся в генах – в последовательности нуклеотидов ДНК. При копировании одной из цепочек ДНК формируется информационная РНК (иРНК), которая служит своеобразной программой для «сборочного конвейера», состоящего из цепочки рибосом. На этом конвейере собираются огромные неуклюжие молекулы полипептидов (поли- - много), которые в дальнейшем с помощью специальных ферментов – протеаз (отсюда – протеолиз) нарезаются на множество белков и пептидов, уже готовых к выполнению своих функций. Это нарезание идет не случайным образом, а между строго определенными парами аминокислот (их называют фланкирующими). Поэтому протеолиз идет не хаотично, а строго прицельно. В результате получаются самые разнообразные пептиды. Покажу вам 3 типичных примера. То, что пептиды могут играть роль и гормонов, и медиаторов, стало известно более полувека тому назад – в 60-х годах прошлого века, благодаря работам француза Роже Гиймена и поляка Анджея Виктора Шалли (став американцами, а в 1977 году – и Нобелевскими лауреатами, они стали называться Роджером Гиллемином и Эндрю Шелли). За прошедшие десятилетия количество пептидов с медиаторными функциями постоянно увеличивалось и сегодня превышает 3 тысячи. Объединены они в более чем 50 семейств. Естественно, я не собираюсь мучить вас всем этим изобилием. Мы обсудим только самые интересные экземпляры. Гипоталамические рилизинг-гормоны Первыми из открытых нейроактивных пептидов были тиролиберин и кортиколиберин, относящиеся к семейству гипоталамических рилизинг-гормонов. Здесь нам придется кратко коснуться связи Роджер Гиллемин и Анджей Шалли нервной и эндокринной систем. Как вы, вероятно, помните, главной эндокринной железой принято считать гипофиз. Это верно только отчасти. У гипофиза есть свой «начальник». Это уже знакомый нам гипоталамус. Гипоталамус управляет гипофизом двумя путями. Первый – сугубо гормональный: от гипоталамуса к передней доле гипофиза (аденогипофизу) тянутся очень короткие и широкопросветные сосуды, по которым в переднюю долю гипофиза поступают рилизинг-гормоны (release - выпуск). Их довольно много – более десятка – и их принято подразделять на либерины и статины. Про второй путь поговорим в разделе «вазитоцины». Либерины стимулируют выработку гипофизом соответствующих тропных гормонов, а статины – тормозят. Гипофиз вырабатывает различные тропины, которые с током крови доносятся до желез третьего порядка: надпочечников, печени, щитовидной железы и др. А уже эти железы выделяют в кровь свои специализированные гормоны (кортикостероиды, соматомедины, тироксин, трийодтиронин и т.д.), которые регулируют обмен веществ в соответствующих клетках-мишенях, у которых есть рецепторы к этим конкретным гормонам. НЕКОТОРЫЕ РИЛИЗИНГ-ГОРМОНЫ ЛИБЕРИНЫ МИШЕНИ В СТАТИНЫ (активация) ГИПОФИЗЕ (торможение) соматолиберин соматотропин соматостатин меланолиберин меланотропин меланостатин пролактолиберин пролактин пролактостатин кортиколиберин АКТГ тиролиберин тиреотропин люлиберин лютропин фоллиберин фоллитропин То, что либеринов намного больше, чем статинов, не должно вас удивлять. Просто там, где активность всей гормональной цепочки (принято называть – эндокринной оси) может регулироваться по отрицательной обратной связи. То есть, при получении нужного количества конечных продуктов оси – каких-то нужных организму молекул – эти молекулы тормозят активность гипоталамуса. Это все касается гормональной регуляции функций нашего организма, но мы ведь обсуждаем медиаторные свойства пептидов. Сейчас я постараюсь вам показать, что они взаимосвязаны. Например, такой рилизинг-гормон, как соматостатин тормозит выработку адендогипофизом соматотропного гормона (или, в просторечии, гормона роста). У соматостатина в паре – соматолиберин. Как ясно из названия, он, наоборот, стимулирует выработку гормона роста. Возможно, вы помните фотографию из школьного учебника, где рядом с ногой баскетбольного центрового стоит лилипут. Только подпись к этой фотографии – вранье! Не в гипофизе там дело, а как раз в гипоталамических рилизинг-гормонах. И мини-пиги, и мини-пони – все о том же. И шестидесятикилограммовые белые крысы (была выращена такая парочка на заре генной инженерии) – тоже о гипоталамусе. А вот как медиатор соматостатин тормозит все, до чего только дотянется: во всех нейронных сетях он выступает как сугубо тормозный медиатор. Другой типичный пример – кортиколиберин. Перед вами – ось стресса. В ней кортиколиберин, высвобождающийся из гипотоламуса при стрессе (кстати, это высвобождение активируется норадреналином, то есть САС), запускает целый эндокринный каскад. Из аденогипофиза в ответ выделяется АКТГ (адренокортикотропный гормон), который с током крови доплывает до коры надпочечников. Кора надпочечников, в свою очередь, выделяются в кровь кортикостероиды, которые в клетках-мишенях стимулируют ионный обмен (это делают минералокортикоиды) и углеводный обмен (глюкокортикоиды). В результате организм получает дополнительные энергоресурсы для активной защиты от повреждающего воздействия. Но это гормональная активность кортиколиберина. А как медиатор центральной нервной системы он обеспечивает целый букет эффектов, очень своевременных в ситуации стресса. Кортиколиберин стимулирует двигательную активность, поисковое поведение (то есть, поведение, в условиях стресса направленное на поиск возможности выхода из критической ситуации) и систему бодрствования. Действительно, в ситуации, когда тебе угрожает опасность, совершенно несвоевременно сладко спать, равно как и заниматься добыванием пищи и продолжением рода. А ведь эти две инстинктивные программы кортиколиберин тормозит. Помните, как адреналин и норадреналин обеспечивают координацию усилий организма на первой стадии стресса? – Кортиколиберин тоже демонстрирует способность формировать согласованные усилия для активной защиты от угрозы на второй стадии (стадии резистентности) стресса. Причем это проявляется и на нейрофизиологическом, и на эндокринологическом, и на энергетическом, и на психическом, и на поведенческом уровнях. Вазотоцины: окситоцин и вазопрессин Это, сравнительно небольшое, семейство медиаторов объединяет нейропептиды, длина молекул у которых не превышает 10 аминокислотных остатков. Эти пептиды обеспечивают второй путь, связывающий гипоталамус с гипофизом – уже не через кровь, а с помощью аксонального транспорта. Мы привыкли к тому, что главной задачей нейрона является проведение нервных импульсов по мембране этого нейрона. Но нейрон – вполне банальная живая клетка, под мембраной которой находится цитоплазма с плавающими в ней органоидами (ядром, митохондриями, рибосомами и т.д.). И сама цитоплазма постоянно перемещается внутри клетки. В середине ХХ века удалось доказать, что это перемещение – постоянное и закономерное (подобно морским течениям, вроде Гольфстрима), и есть целый ряд молекул – прежде всего, пептидов – которые с помощью этого тока цитоплазмы переносятся от ядра клетки к пресинапсам. То есть, аксоны (а иногда и дендриты) в этом случае выступают в качестве своеобразных трубопроводов. Именно таким способом вазотоцины, синтезирующиеся в нейронах гипоталамуса, доставляются в заднюю долю гипофиза (нейрогипофиз), где и накапливаются, чтобы потом выделиться в общий кровоток. Мы коснемся двух вазотоцинов. Окситоцин часто называют «гормоном молодой мамы». И снова надо признать, что прозвище очень далеко от истинного портрета вещества. Вопервых, это не только гормон, но и медиатор. Во-вторых, его активность особенно возрастает до родов, когда женщина еще только готовится стать мамой. Наконец, возраст будущей мамы не имеет никакого значения. Эффекты, благодаря которым исследование окситоцина до сих пор остается актуальной задачей, действительно, связаны с подготовкой организма матери к рождению ребенка. Окситоцин стимулирует ритмические сокращения мускулатуры матки – те самые родовые схватки. Он же активирует лактацию, то есть выработку молока. Действительно, когда ребенок родится, его хорошо бы покормить. У окситоцина есть еще несколько эффектов, но нам интереснее всего его способность угнетать память. Именно его действием во многом объясняется несколько наивное поведение беременных и недавно ставших мамами женщин. Есть гуманистическая гипотеза, согласно которой окситоцин как молекулярный ластик стирает часть воспоминаний роженицы, чтобы та излишне не концентрировалась на воспоминаниях об очень болезненной процедуре родов. Но это – только гипотеза… Вазопрессин отличается от окситоцина всего парой аминокислот. Но эффекты, которые он вызывает, существенно другие. Прежде всего, вазопрессин оказался Естественно, на очень основе этого эффективным его стимулятором свойства попытались памяти. создать лекарственный препарат для укрепления памяти (мечта студента, не правда ли?). Однако дальше экспериментов на животных дело не пошло. Почему? – Помешали другие эффекты вазопрессина. Прежде всего, само его название – «говорящая фамилия». «Вазо-» - сосуды, речь идет о кровеносных сосудах, «пресс» - и без перевода понятно, что речь идет о сжатии. То есть вазопрессин способен сжимать кровеносные сосуды, повышая таким образом кровяное давление. Едва ли найдутся желающие ради улучшения памяти рисковать получить инсульт или инфаркт. Но у вазопрессина есть и еще одно название – антидиуретический гормон (АДГ). Дело в том, что для успешного выведения из организма азотсодержащих ядов (NH?, NO?) необходимо, чтобы давление крови в так называемых Мальпигиевых клубочках (сосудистых сплетениях, из которых жидкая составляющая крови выжимается в капсулы Шумлянского – важнейшие начальные элементы нефронов наших почек) было высоким. Это «прессование» крови, как мы уже выяснили, способен обеспечить вазопрессин. Но при этом образуется непомерно большое количество очень жидкой мочи (первичной мочи). Если бы на этом работа почек заканчивалась, жизнь животных была бы чрезвычайно однообразной: целыми сутками они занимались бы только тем, что пили и мочились. Но природа гуманна и разумна. Первичная моча проходит в нефронах через извитые каналы (петли Генле), где бОльшая часть воды с растворенными в ней солями возвращается в кровь, а «на выход» отправляется концентрированная вторичная моча – биологическое оружие массового поражения. Я не шучу: во вторичной моче так высока концентрация ядов, что шансов выжить у попавших под струю растений или животных остается очень мало. Не зря в войну на поле боя смоченные мочой повязки использовали для защиты ран от заражения. Но вернемся к вазопрессину. Именно он стимулирует в почках возвращение воды в кровь, снижая, таким образом, диурез (мочеотделение). Отсюда и название – антидиуретический. Меланокортины: АКТГ и МСГ К следующему семейству нейропептидов относятся гормоны аденогипофиза и, одновременно, медиаторы: адренокортикотропный гормон (АКТГ) и меланоцитстимулирующий гормон (МСГ). Здесь нам интересен АКТГ. Как гормон, он включен в уже обсуждавшуюся нами ось стресса: выделяется из гипофиза в ответ на сигнал кортиколиберина и с током крови достигает своих рецепторов в коре надпочечников, стимулируя высвобождение кортикостероидов. Но, оказывается, что у АКТГ есть очень важное медиаторное свойство: он является мощным стимулятором памяти (точнее, запоминания). Причем, если активировать энергетический обмен (катаболизм) при стрессе умеет только цельная молекула АКТГ (в ней 39 аминокислотных остатков – один из самых длинных нейропептидов), то стимулировать запоминание может не только вся молекула, но и ее фрагменты: АКТГ 4-7, АКТГ4-10 и АКТГ1-24. Это открытие позволило в СССР создать эффективный лекарственный препарат Семакс (АКТГ4-7-Про-Гли-Про; то есть 4 аминокислоты из АКТГ, к которым присоединены пролин, глицин и еще один пролин, как защита от ферментов), который нашел применение в психоневрологических клиниках всего мира. Только хочу предупредить: это лекарство для больных, а не биодобавка для студентов, мечтающих за последнюю ночь перед экзаменом выучить все билеты! Брадикинины и тахикинины: брадикинин, субстанции Р Нетрудно заметить, что в названиях обоих семейств коренным словом является «-кинины». Это связано с тем, что одним из способов выявления медиаторных свойств пептидов является проверка его действия на гладкие мышцы внутренних органов. Это могут быть полоски, вырезанные и стенки аорты или тонкого кишечника, фрагменты мышцы сердца (в сердце у нас некий гибрид поперечнополосатой и гладкой мускулатуры) и т.д. В лаборатории проверяют, вызывает ли изучаемый пептид сокращение таких препаратов, и как происходит сокращение: быстро или медленно. При тестировании пептидов этих двух семейств выяснилось, что одни из них вызывают медленное сокращение гладкой мускулатуры; они получили название брадикининов (бради- - медленно: брадикардия – редкий пульс, брадипноэ – редкое дыхание, и т.п.). Другие вызывали быстрое сокращение, и стали называться тахикининами (сравните: тахикардия – это когда сердце колотится, например, после быстрого бега). Брадикинины вызывают в нашем организме комплекс эффектов, который легче всего запомнить, если вспомнить те ощущения, которые вызывает ужаление осы. Я не шучу: главной составляющей осиного яда является именно брадикинин. Отсюда – типичные проявления: расширение кровеносных сосудов (покраснение в месте ужаления), снижение кровяного давления, отек, боль, затруднение дыхания, усиление агрессии. Особого внимания заслуживают тахикинины. Они тоже обнаружены у самых разных видов животных: у моллюсков, насекомых, многоножек и т.д. Но у млекопитающих (то есть и у нас с вами) типичным тахикинином является субстанция Р (Р – латинское). Надо сказать, что ее открытие является поучительным примером когнитивной слепоты, когда давлеющее мнение социума (в данном случае – научного) мешает увидеть очевидное даже выдающимся умам. В 1931 году два известнейших биохимика и фармаколога того времени швед Ульф фон Эйлер и англичанин Джон Гаддум сообщили об обнаружении нового – второго после ацетилхолина – медиатора с целым набором своеобразных свойств. Они назвали это вещество «субстанцией Р», потому что при консервировании его с помощью лиофильной сушки оно приобретало вид мельчайшей пудры (powder). Однако затем у исследователей возникли так и не преодоленные в итоге трудности. Для того чтобы с точностью охарактеризовать химическую структуру биологически активного вещества, надо его очистить от примесей, в том числе – белковых. Проверка на чистоту включает доказательство отсутствия в исследуемом растворе пептидных связей, то есть тех ниточек, на которые нанизаны бусинки аминокислот в белках и пептидах. Методика простая, рутинная: окрасился раствор – есть «ниточки», остался бесцветным – раствор очистили от белков. И вот на этом этапе начались сплошные неприятности. Когда раствор с субстанцией Р очищали от предполагаемых белков, он терял активность и, соответственно, медиаторные свойства. предыдущей, окрашивающейся Как только возвращались реактивом порции, к активность восстанавливалась во всей красе. И так раз за разом, год за годом. В соответствии с тогдашним научным «общественным мнением», белки категорически не могли быть медиаторами, слишком они громоздки и тяжелы (замечу осторожно, что с этим и сегодня спорить не приходится). А вот про пептиды в то время никто и подумать не мог, хотя их молекулы не намного больше классических медиаторов, а пептидных связей, вызывающих окрашивание проверяемого раствора, в них вполне достаточно. Итог: в соответствии с принципами научной добросовестности, фон Эйлер и Гаддум в 1937 году опубликовали статью, в которой покаялись в совершенной, по их мнению, методической ошибке и «закрыли» собственное открытие субстанции Р. Забавно, что когда уже в 70-х годах ХХ века субстанция Р была открыта заново и обозвана сложным набором букв и цифр (фантазия на имена собственные для пептидов начала уже иссякать), доживший до этого момента уже совсем пожилой фон Эйлер опознал свое брошенное полвека назад детище и вернул ему первоначальное имя. Курьёзно, что субстанция Р в дальнейшем продолжала оставаться «пептидным бесёнком». В 80-х годах ХХ века из-за неё чуть ли не напрочь перессорились специалисты по физиологии боли и обезболивания. Одни лаборатории сообщали о выраженных болеутоляющих свойствах пептида, а другие – о том, что субстанция Р вызывает боль. Дело дошло до обоюдных обвинений в некомпетентности. Примирить позиции спорщиков удалось на одном из конгрессов, когда выяснилось, что всё зависит от того, куда вводится вещество: в кровь или сразу в мозг. Один и тот же пептид, в зависимости от способа введения, вызывает прямо противоположные реакции. – Какая неожиданность! Кто мог тогда предполагать, что регуляторные пептиды ещё не один раз будут преподносить такие сюрпризы. Опиоидные пептиды: эндорфины, энкефалины, динорфины и др. Наконец, последнее семейство из тех, которые мы выбрали для обсуждения: опиоидные пептиды. Название пептидов этого обширного семейства объясняется следующим. В медицине давно и хорошо известна группа так называемых наркотических анальгетиков, то есть очень эффективных обезболивающих лекарств, вызывающих, к несчастью, смертельную и почти неизлечимую болезнь – наркоманию. Эти лекарства называются опиатами, потому что начало этой группе препаратов послужил экстракт сока опийного мака – опий (или опиум). К этой группе относятся промедол, морфин, фентанил, героин, пентазоцин, метадон и мн.др. Суффикс «-оид-» подобие. Вспомните, в клетке есть органоиды: рибосомы, ЭПР, ядро и др. Неприлично было бы назвать их органами, ведь органы состоят из клеток (получится сказка про белого бычка), поэтому: органоиды, то есть подобные органам. Стало быть, опиоиды – это вещества (прежде всего, пептиды), по своим свойствам подобные опиатам. История их открытия чрезвычайно драматична и заслуживает отдельного разговора (вот доучитесь до третьего курса – тогда услышите). Главное, что в 70-х годах прошлого века сначала были открыты мембранные рецепторы к опиатам, названные опиатными рецепторами (ОР). Оказалось, что их как минимум 3 типа: μ- (мю-), δ- (дельта-) и χ- (каппа-). Буквально спустя полтора года были обнаружены первые эндогенные (то есть, напомню, внутренние) лиганды к этим рецепторам: пептиды β-эндорфин, мет-энкефалин и лей-энкефалин. За прошедшие с тех пор почти полвека таких эндогенных опиоидных пептидов в нашем организме обнаружили уже почти 50. В таблице приведены только некоторые из них и их структура в однобуквенном аминокислотном коде. Не подумайте плохого: это пример, его не надо заучивать! Оказалось, что биологические свойства большинства из опиоидов очень близки свойствам опиатов. Они вызывают эйфорию, равнодушие к проблемам, торможение работы внутренних органов, обезболивают и т.д. В целом эффекты опиоидных пептидов минимизирующие и гипобиотические. можно охарактеризовать как Что это значит? Эндогенные опиоиды снижают активность большинства систем нашего организма при ее чрезмерном усилении – это минимизация. Кроме того, они могут перевести организм в состояние крайней экономии энергоресурсов – это гипобиоз. Примером гипобиоза является гибернация (то есть зимняя спячка) наших местных летучих мышей, лягушек или ежиков. У них температура тела падает больше, чем на 20 градусов, резко замедляется пульс, урежается дыхание, перестает работать пищеварительная система, и др. Для человека - это крайний случай, например, летаргический сон. Но в более мягкой форме это проявляется при шоке и на третьей, последней стадии стресса (стадии истощения). Вообще роль эндогенной опиоидной системы (ЭОС: она объединяет опиоидные пептиды и опиатные рецепторы) при стрессе очень важна. Она ограничивает избыточную активацию САС на первой стадии, продолжает свою деятельность на второй стадии и является полновластной хозяйкой третьей стадии. Наверное, вы испытывали бессилие и вялость после перенесенного сильного волнения. Эта работа ЭОС, и направлена она на вашу защиту (только теперь уже пассивную), то есть экономию и восстановление ваших сил. К концу ХХ века выяснилось, что ЭОС является самой древней на нашей планете регуляторной системой: она обнаружена у всех живых существ на Земле. Ее собратья по управлению стрессом – САС и ГГАС – несравнимо, на многие миллионы лет моложе. Естественно, открытие эндогенных опиоидных пептидов позволило внести ясность в механизмы развития страшной болезни – опиатной наркомании. Оказалось, что опиатные рецепторы очищаются от отработавших опиоидов путем интернализации (надеюсь, вы еще не забыли этот механизм?). Когда вместо родного опиоида опиатному рецептору подсовывают опиатный наркотик, сам рецептор обмануть удается, и в организме развиваются те же эффекты, что и под влияние опиоидов (например, обезболивание). Но обмануть процесс интернализации невозможно, потому что ферменты пептидазы, расщепляющие опиоидные пептиды до отдельных аминокислот, ничего не могут сделать с алкалоидами, каковыми являются опиатные наркотики. Нет в молекулах алкалоидных наркотиков пептидных связей, которые умеют расщеплять пептидазы. В результате рецептор не очищается, вернуться на мембрану не может и гибнет. Поэтому у наркомана с каждым приемом наркотика количество опиатных рецепторов уменьшается, и ему приходится увеличивать дозу наркотика, чтобы добиться того же эффекта. Развивается толерантность, то есть нечувствительность к прежней дозе. И так – шаг за шагом, пока передозировка наркотика не приведет или к остановке сердца и дыхания, или к перитониту (воспалению забрюшинного пространства), или к выключению почек, или к разрушению иммунной системы (СПИД у наркоманов – закономерный результат развития толерантности). В любом случае финиш такой жизни достигается достаточно быстро и мучительно. К сожалению, в некоторых случаях люди становятся наркоманами не по собственной глупости, а в связи с введением наркотических анальгетиков по медицинским показаниям: например, при необходимости проведения серьезной операции или для обезболивания при невыносимых болях. Это так называемая ятрогенная (вызванная медицинским вмешательством) наркомания. Так что нельзя всех наркоманов стричь под одну гребенку. Да и вообще, самый опустившийся наркоман – все равно человек. Поэтому уже не первый век идет поиск эффективных анальгетиков, не вызывающих наркомании. Собственно, вся линейка опиатных анальгетиков (даже героин!) была создана в связи с этим поиском, да и открытие ЭОС тоже результат этого поиска. И когда были обнаружены опиоидные пептиды, появилась надежда, что на их основе удастся создать такие болеутоляющие средства, которые станут обезболивать не хуже наркотических анальгетиков, но не будут вызывать наркомании. Попыток было много, но пока успех не достигнут. Например, большие надежды возлагались на препарат даларгин (d-Ала2-Лей5-энкефалин, то есть лей-энкефалин, в котором находящийся на втором месте глицин заменен на правовращающий изомер аланина). Оказалось, что его болеутоляющая активность никуда не годится, но с его помощью можно успешно лечить язвенную болезнь желудка и тонкого кишечника. И вдруг в 2020 году выяснилось, что даларгин может помочь в лечении короновирусной инфекции. Вот уж не знаешь: где найдешь, где потеряешь. Сегодня ясно, что ЭОС участвует во всех сферах нашей физической и психической жизни. Поэтому напоследок хочу вспомнить пророчество уже знакомого вам Нобелевского лауреата Роджера Гиллемина (он же - Роже Гиймен), который почти 50 лет назад утверждал, что открытие эндогенных опиоидных пептидов приведет к пересмотру наших знаний и в области наркологии, и в реаниматологии (наука об оживлении), и в психиатрии, и в представлениях о научении, памяти, эмоциях, мотивациях, мышлении и даже эволюции. Я хорошо помню, как особо жизнерадостные студенты буквально умирали от хохота, когда я им об этом тогда рассказывал. А ведь надо же – все сбылось! ВТОРЫЕ МЕССЕНДЖЕРЫ Как вы уже знаете, кроме медиаторов, к информонам относятся вторые мессенджеры (или, как неграмотно и, к сожалению, часто их называют – вторичные посредники). В чем, казалось бы, разница? – Разница существенная. Вторичным бывает сырье, например, макулатура или ржавые кастрюли. Вторичными становятся продукты, после того как мы их съели. А мессенджеры – именно вторые по порядку. Первооткрыватель этой группы информонов американский биохимик и фармаколог Эрл Сазерленд-младший, закономерно получивший в 1971 году Нобелевскую премию, рассуждал просто и понятно. Многие гормоны и медиаторы доносят информацию до мембраны клетки – и их нужно считать первыми (по порядку) передатчиками сигналов. А дальше – как эстафетная палочка – информация передается на систему внутриклеточных передатчиков. По порядку, по счету они вторые (second messenger). Итак, вторые мессенджеры – это внутриклеточные передатчики информации. Они высвобождаются в ответ на активацию рецепторов и транслируют сигнал внутри клетки. Именно благодаря этому реализуется «медленный способ» активации лигандзависимых ионных каналов постсинаптической мембраны. Но вторые мессенджеры обеспечивают трансляцию информации в клетки не только в нервной системе. Многие гормоны (например, производные аминокислот, и белково-пептидные гормоны), не способные проникать в клетку через ее мембрану, управляют обменом веществ в клетке именно с помощью вторых мессенджеров. Особенностью активности вторых мессенджеров является их способность запускать внутри клетки различные каскады биохимических реакций, которые в итоге приводят к управлению такими важнейшими для организма процессами, как пролиферация клеток, их дифференцировка, миграция или апоптоз. Подозреваю, что каждое из этих слов требует объяснения. Пролиферация – это деление клеток (вспомните школьное – митоз). Клетки нашего тела обновляются непрерывно, и происходит это именно благодаря делению. Да и с самого начала – от оплодотворенной яйцеклетки – вся наша жизнь основывается на постоянном делении клеток. Дифференцировка – приобретение исходно одинаковыми клетками новых, существенно различающихся свойств – нужна тоже не только на начальных этапах онтогенеза (там-то это естественно и понятно: именно благодаря дифференцировке у нашего зародыша появляются и нервная, и покровная, и мышечная, и другие специализированные ткани). Все вы слышали про стволовые клетки. Они есть в каждой из систем нашего организма. Это клетки, дифференцироваться в которые различные исходно узко обладают способностью специализированные группы. Например, предшественниками наших главных иммунных защитников – лейкоцитов – являются такие стволовые клетки, которые в результате «воспитания» (не только вас воспитывают, клетки тоже!) порождают и Ткиллеров, и Т-хелперов, и Т-супрессоров, и В-лимфоциты. А это уже жестко дифференцированные бойцы нашего невидимого фронта, способные на корню задушить почти любую инфекцию. Ненавистный короновирус ухитряется их обмануть только потому, что у него есть СПИДообразные свойства, позволяющие успешно маскироваться. Зачем нужна миграция клеток? – А это тоже этапы нашего онтогенеза. Например, сетчатка наших глаз образуется в результате миграции нейронов промежуточного мозга. То есть, сетчатка – это «мозги на выселках». Из того же промежуточного мозга путем миграции образуется задняя доля гипофиза (нейрогипофиз) – главной эндокринной железы (вспомните гипоталамические рилизинг-гормоны). нашего организма Наконец, что такое апоптоз? – А это запрограммированное самоубийство клеток. Когда клетка исчерпывает ресурсы своей активности и становится дармоедом, или когда в ней произошли такие нарушения, что она начинает вредить организму, в котором она живет, запускается генетически детерминированный механизм ее самоуничтожения. Важно, чтобы ядовитые продукты ее распада не навредили окружающим здоровым клеткам. Поэтому программа апоптоза строго отрегулирована. Одна из популярных гипотез, объясняющих возникновение рака (канцерогенез), считает сбой в программе апоптоза одной из главных причин этой страшной болезни. Всеми этими процессами умеют управлять вторые мессенджеры. Кроме того, именно с их помощью можно заставить клетку усилить или анаболизм (пластический – строительный – обмен), или катаболизм (энергетический обмен). Не случайно рецепторы, активация которых приводит к запуску системы вторых мессенджеров, называются метаботропными. И, наконец, мы с вами уже обсуждали, что вторые мессенжеры действуют через каскад активации протеинкиназ – ферментов, способных модифицировать структуру белков путем их фосфорилирования, то есть присоединения к аминокислотам этих белков остатков фосфорной кислоты (опять же, вспомните: в АТФ целых три таких остатка). В результате меняется внутренняя энергия такого белка, изменение внутренней энергии ведет к изменению конформации белковой молекулы (пружина сжатая и пружина растянутая), а это, в свою очередь, ведет к ее активации. Если это, например, фермент, он переходит в активное состояние и начинает катализировать реакции, за которые он отвечает. Если это белок ионного канала, изменение его конформации ведет к открыванию канала, и в открытом состоянии, как мы уже обсуждали, такой канал может находиться долго. Насколько важны протеинкиназы, можно судить по тому, что их кодированием занимается примерно 2% нашего генома. Думаю, не нужно больше убеждать вас в необходимости вторых мессенджеров для работы нашего организма. Пришла пора поближе познакомиться с некоторыми из них, благо, пока мы знаем их не так уж много. Циклические нуклеотиды (цАМФ и цГМФ) Прежде всего, почему нуклеотиды. Надеюсь, вы еще не забыли, как учили в школе А, Т, Ц, Г – четыре нуклеотида (аденин, тимин, цитозин, гуанин), которые формируют ДНК. Но это не единственная функция нуклеотидов. Здесь мы увидим, как они участвуют во внутриклеточной передаче информации. Аденозинтрифосфат – АТФ Циклический аденозинмонофосфат – цАМФ Первым из вторых мессенджеров (извините за оксюморон!), открытым Сазерлендом, был циклический аденозинмонофосфат (цАМФ). Формулы я привел вовсе не из садистских побуждений: я хочу, чтобы вы увидели родство второго мессенджера и главного энергетического аккумулятора наших клеток. Действительно, цАМФ образуется из АТФ, а катализирует эту реакцию фермент аденилатциклаза. А этот фермент, в свою очередь, активируется «по приказу» рецептора через систему так называемых Gбелков. В результате цАМФ транслирует сигналы, переданные многими медиаторами на рецепторы, и на ионные каналы постсинаптической мембраны, и на внутриклеточные протеинкиназные каскады. Посмотрите, например, как этот циклический нуклеотид реализует эффекты уже известных вам опиоидных пептидов: Активность опиоидных пептидов реализуется через метаботропные опиатные рецепторы. Внутриклеточные каскады запускаются с помощью цАМФ (cAMP). Активировать цАМФ можно, например, всем известным кофеином. Именно поэтому любители кофе и чая (формально в чае кофеина даже больше, чем в кофе) используют эти напитки для улучшения когнитивных функций. Но, как и положено допингу, все хорошо в меру. Превышение оптимальных для каждого концентраций кофеина (и других производных ксантина) может привести к ингибированию активности цАМФ и, следовательно, прямо противоположному эффекту. Передозировка кофеина может вызвать сонливость, что нужно учитывать в последнюю ночь перед экзаменом. Циклический гуанозинмонофосфат (цГМФ) тоже является результатом отщепления двух фосфорных остатков гуанозинтрифосфата катализируется Циклический гуанозинмонофосфат – цГМФ (ГТФ). ферментом от исходного Эта реакция гуанилатциклазой. Обратите внимание на «говорящие фамилии» этих нуклеотидов: добавляем к азотистому основанию аденину сахар – получается аденозин, добавляем сахар к азотистому основанию гуанину – получается гуанозин. Долгое время цГМФ приписывали свойства, прямо противоположные цАМФ. Сегодня стало ясно, что у этого второго мессенджера есть вполне самостоятельные функции. Например, цГМФ регулирует электролитный обмен, то есть обмен ионов и обмен воды. Кроме того, цГМФ участвует в релаксирующем действии на сердечную мышцу еще одного второго мессенджера – оксида азота (NO), о котором речь у нас еще впереди. Липиды мембраны (инозитолтрифосфат и диацилглицерол) Не удивляйтесь: фосфолипиды мембраны не только образуют активную пленку, окружающую цитоплазму клетки. Они сами – точнее, продукты их расщепления – могут проявлять самостоятельную активность, выступая в роли вторых мессенджеров. И главными среди них являются инозитолтрифосфат и диацилглицерол, которые образуются при гидролизе (то есть расщеплении при взаимодействии с водой) мембранного фосфолипида фосфоинозитолдифосфата с помощью фермента фосфолипазы С. Свои внутриклеточные эффекты эти липиды, в основном, реализуют через регуляцию обмена ионов Ca2+ и активацию протеинкиназных каскадов. Схематическое представление об этом вы можете увидеть на схеме ниже: Кальмодулин Кальмодулин относится к семейству Ca2+связывающих белков. Вы уже встречались с подобными тропонин). (вспомните Он был тропомиозин открыт и сравнительно недавно – в 1980 году. Оказалось, что он тоже может выступать в роли второго мессенджера, Молекулярная структура Ca -связывающего белка 2+ кальмодулина обеспечивая активацию целого ряда внутриклеточных ферментов. Я не случайно привел здесь его молекулярную структуру. Она позволяет представить его кальций- связывающий механизм. Два полярных участка молекулы, каждый из которых имеет по два сайта для связывания Ca2+, связаны центральной шарнирной спиралью. В отсутствии Ca2+ белок не активен, но при связывании с кальцием меняет свою конформацию и переходит в активное состояние. Помните, мы обсуждали, что ионы кальция – самые активные ионы в организме. Вот вам еще одно подтверждение. Оксид азота (NO) В 1998 году американским ученым Роберту Форшготту, Ферид Мьюрэд и Луису Игнарро была вручена Нобелевская премия по физиологии и медицине с такой формулировкой: «Нобелевская премия по физиологии и медицине присуждена за открытие роли оксида азота как сигнальной молекулы в сердечнососудистой системе». В разъяснении формулировки указывалось, что этим ученым удалось выяснить, почему такое опасное взрывчатое вещество как нитроглицерин в малых дозах способно спасти жизнь больным-сердечникам при остром приступе стенокардии (сто лет назад эта болезнь называлась «грудная жаба»). Действительно, Форшготт, Мьюрэд и Игнарро проследили цепочку превращений нитроглицерина в организме и обнаружили, что от него отщепляется такая крохотная молекула как NO, и она способствует снятию спазма коронарных (внутрисердечных) сосудов, предотвращая гипоксию (недостаток кислорода) и ишемию сердечной мышцы – миокарда. При исследовании механизма такого спасительного действия оксида азота обнаружилось, что он является и своеобразным медиатором, и, что для нас сейчас важно, вторым мессенджером. Нужно сказать, что это открытие немного смягчило негативное, в целом, отношений специалистов самых разных областей знаний к азотсодержащим малым молекулам (веселящий газ, отравление атмосферы Земли, Грета Тумберг и прочие неприятные последствия). Другие вторые мессенджеры К сожалению, наш короткий курс не позволяет глубже окунуться в эту интереснейшую тему. Хочу только сказать вам, что каждое новое открытие в области вторых мессенджеров расширяет наши знания об устройстве внутриклеточной кухни нашего организма. За последние десятилетия открыты такие вторые мессенджеры как СО (угарный газ!), c-jun, c-fos и др. Оказалось, что в результате расшифровки особенностей их действия мы существенно продвинулись и в понимании механизмов памяти (здесь большая роль принадлежит отечественному нейрофизиологу академику К.В. Анохину), и в объяснении двигателей канцерогенеза (развития раковых опухолей), и в причинах сердечнососудистых заболеваний… Но это уже другая история. НЕЙРОНАЛЬНЫЕ МОДУЛИ или МАЛЫЕ НЕЙРОННЫЕ СЕТИ До сих пор мы с вами говорили или о работе одного нейрона, или о взаимодействии двух связанных синаптически нейронов. Но ведь мозг – это множество сложнейших нейронных сетей. Следовательно, нужно выяснить, есть ли какие-нибудь общие принципы, правила функционирования таких сетей. В начале второй половины ХХ века появились 2 книги, то время которые в произвели революцию в Карл Х. Вернон Б. Джералд М. умах Прибрам Маунткасл Эдельман «Языки мозга» К.Прибрама нейрофизиологов: и «Разумный мозг» В.Маунткасла и Д.Эдельмана. В них впервые был поставлен вопрос о необходимости поиска закономерностей работы нейронных сетей и вполне справедливо предлагалось начинать это с относительно простых образований – нейрональных модулей. Еще тогда было понятно, что такие принципы должны быть универсальными: проявляться как в ганглиях примитивных животных (например, виноградной улитки), так и в огромном человеческом мозгу. И работа закипела! Целые группы нейробиологов стали работать с ганглиям виноградных улиток или даже примитивной нематоды (помните таких круглых червей из предыдущего уровня?) Caenorhabditis elegans (C. elegans). Caenorhabditis elegans Параллельно велся поиск тех же принципов в мозгу гораздо более совершенных животных, в том числе млекопитающих и даже приматов. А еще одно направление было напрямую не связано с живыми существами – это математическое моделирование процессов в нейронных сетях. Как раз за такие исследования теперь уже знакомый вам Джеральд Эдельман получил Нобелевскую премию. Курьезно, что при этом было нарушено завещание Альфреда Нобеля, категорически запретившего присуждать его премию уже умершим и математикам. Умершим, потому что Нобель предполагал, что его оставленное ученым Мира наследство поможет им продолжить важные исследования, не заботясь о куске хлеба. Стало быть, премию надо давать живым и молодым. Сам А. Нобель на своей шкуре испытал все «удовольствия» научной деятельности, пока не стал работать в чисто военной науке. Парадокс: если ты изобрел способ спасения жизней 100 человек, тебя, в лучшем случае, похвалят. А вот если ты придумал, как тех же 100 человек в одночасье отправить в мир иной, ты – миллионер. А вот второй запрет, судя по легенде, связан с сугубо личными проблемами Нобеля: якобы какой-то математик «увел» у него жену. Вот такая страшная месть! На самом деле, Альфред Нобель никогда не был женат. Так что это легенда. Известно однако, что он жестко конфликтовал с выдающимся шведским математиком Магнусом Миттаг-Леффлером и очень неприязненно отзывался обо всех математиках вообще. Все может быть… Но вернемся к Д. Эдельману. Чтобы обойти запрет Нобеля, Эдельмана специалиста в области математического моделирования - назвали иммунологом, и на этом успокоились. Как бы то ни было, но к концу ХХ века основные принципы функционирования нейрональных модулей были выяснены. Прежде всего, договорились, что такое нейрональные модули или малые нейронные сети. Это небольшая (от 3 до 100000) группа нейронов, связанных между собой синаптически и выполняющих какую-то одну простейшую вычислительную функцию. Простейшие вычислительные функции модулей Что значит – простейшая вычислительная функция? Их несколько, но нам интересны 5 базовых: - усиление сигнала; - подавление сигнала; - выделение контраста; - поддержание ритмики; - поддержание тонуса. Зачем нужно усиление сигналов в нейронных сетях, я думаю, объяснять не надо. Именно благодаря этой функции мы слышим малейший шорох (особенно, ночью – в темноте и тишине). Именно поэтому мы видим мельчайшие изменения цветов; особенно это свойственно художникам. Именно поэтому собаки обучаются по запаху искать наркотики, взрывчатку и даже короновирус. Сложнее объяснить, зачем нужны модули, подавляющие сигнал. Но если вы вспомните про боль, смысл этой функции станет очевидным. Вспомните опиоидные пептиды. Модули, в которых есть эти опиоиды, как раз такой сугубо гуманистической работой и занимаются. Теперь о выделении контраста. Нетрудно догадаться, что если информацию обрабатывают два модуля с функциями усиления и подавления сигнала, на границе их активностей возникает усиление контраста. Слева вы видите хорошо известные в психофизике полосы Маха. В конце XIX века австрийский физик Эрнст Мах решил с помощью Полосы Маха точных физических проверить, является приборов ли широко известная иллюзия осветления более светлого и затемнения более темного на границе двух поверхностей (одна – темная, другая – светлая) физическим эффектом или сугубо психическим явлением. Взгляните на рисунок. Действительно, кажется, что на границах двух соседних полос возникает усиление контраста (грани даже кажутся выпуклыми). Мах заподозрил, что мы имеем дело с дифракцией или интерференцией лучей. Какого же было его удивление, когда приборы показали, что интенсивность окраски каждой из этих полос внутри полосы абсолютно одинаковая. Единственное объяснение, которое он нашел: иллюзию контраста создает наша психика, то есть наш мозг. Из этого следует очень важный вывод: то, что мы видим – совершенно не обязательно то, что есть на самом деле. Это категорически противоречит постулату теории отражения В.И. Ленина о том, что «материя – это категория для обозначения объективной реальности, данной нам в ощущениях». Эрнст Мах доказал, что ощущения нам всегда подвирают, и Владимир Ильич его за это сильно невзлюбил и взгляды его сторонников обозвал «махизмом». Но возникает вопрос: а зачем наши модули так старательно усиливают контраст? – А просто представьте себе, как вы в сумерках спускаетесь по незнакомой лестнице. Только благодаря усилению контраста между ступеньками и промежутками между ними мы избавляемся от опасности сломать себе шею. Теперь о ритмах. В курсе «Зоописхологии» мы уже говорили о биоритмологии. Действительно, мы – целая часовая переполненная хронометрами. Большие отрезки времени мастерская, отмеряют гормональные часы, а вот маленькие – доли секунд и секунды – навязывают именно нейрональные модули, которые называются пейсмекерами. Они есть и у нас в мозгу, и в сердце, и в других органах. Наконец, поддержание тонуса. Первое, что приходит в голову, это тонус мышц. Про это все слышали. Действительно, для поддержания постоянного тонуса мышц требуются специальные модули, которые периодически активируют мотонейроны, управляющие мышцами. Полностью расслабленная мышца – это совсем не то, что мы ощущаем, отдыхая на диване. Для полного расслабления нужно, чтобы никакие сигналы от мотонейронов до мышц не доходили. Вспоминайте: мы с вами говорили об яде кураре и производных от него препаратах – миорелаксантах. Вот при их действии, когда заблокированы холинорецепторы, и сигнал не передается в нервно-мышечном синапсе, мы увидим, действительно, мышцу без тонуса: она похожа на кисель или студень – слегка толкнешь, и она будет некоторое время колебаться. Но поддержание тонуса (тонической активности) нужно и для нейронов. Помните, «молчащий нейрон – мертвый нейрон»? Если нейрон хотя бы несколько минут не будет проявлять хотя бы минимальную активность, его перестанут снабжать питательными веществами (глюкозой, в первую очередь) и кислородом. И помрет он, бездельник, позорной смертью от голода. Поэтому специальные модули время от времени «прозванивают» связанные с ними нейроны, будя их и напоминая, что надо бы поработать (или, хотя бы, сымитировать работу). Известен еще целый ряд простейших функций нейрональных модулей, но мы их пока касаться не будем. Перейдем к основным принципам их работы. Принципы работы нейрональных модулей Этих принципов тоже довольно много, но мы с вами разберем пять главных. 1. Дивергенция. Это своего рода «ветвление» нейронных сетей. Аксон от нейрона А у окончания ветвится на два окончания, стимулируя таким образом Благодаря нейроны этому В и С. информация рассылается по нескольким (в нашем случае, двум) адресатам. Такого рода модули представлены во всей нервной системе, но наиболее типичным примером являются сенсорные сети, задача которых – «разослать» полученную информацию по возможно большему количеству адресатов. Например, если вы, не дай Бог, укололи палец, сигнал по афферентным нейронным сетям будет направлен и в спинной мозг, и в разные отделы ствола головного мозга, и в мозжечок, и в промежуточный мозг, и даже в кору больших полушарий головного мозга. 2. Конвергенция. Это, нескольких аксонов на 1 нейрон. наоборот, «схождение» окончаний Согласно этому принципу, активация нейрона С может происходить при разных условиях (давайте договоримся, что про торможение _ ТПСП – мы сейчас вспоминать не будем, чтобы не запутаться окончтельно). Например, нейрон С может активироваться, если суммарный ВПСП на его мембране при активации нейрона А будет надпороговым, то есть достаточным для генерации ПД на субсинаптической мембране нейрона С. Активность нейрона В в этом случае не принципиальна. Точно так же при надпороговом значении ВПСП от нейрона В на субсинаптической мембране нейрона С возникнет полноценный ПД, и активность нейрона А не будет нас интересовать. Наконец, возможен вариант, когда ВПСП от обоих входных синапсов будут недостаточны (подпороговыми) для активации нейрона С. Тогда решающую роль будет играть результат суммации ВПСП от обоих входов при синхронной активации нейронов А и В. Если суммарный ВПСП окажется надпороговым, то мы получим полноценный ПД на субсинаптической мембране нейрона С. Эти рассуждения можно выразить очень просто: а) ВПСПА→ПДС, ВПСПВ—x-> ПДC; б) ВПСПВ→ПДС, ВПСПА—x->ПДС; в) ВПСПА—x->ПДС; ВПСПВ—x-> ПДC; ∑ВПСПА+В→ПДС. Напоминаю, что ∑ - это знак суммы. Не думайте, что я поставил себе задачу свести вас с ума накануне сессии. Эти рассуждения нам понадобятся для рассмотрения двух следующих принципов. Но нам нужно еще разобраться, где используется конвергенция. И снова ответ: везде, во всей нервной системе. Но есть особенно яркий пример. Вспомните мотонейроны спинного мозга, задача которых управлять работой мышц. Управлять очень просто, двоично: пошлет мотонейрон импульсы к мышечным волокнам – те сократятся, не пошлет – не сократятся (все по приказу: дисциплина!). Как здорово, просто мечта любого ефрейтора. Только жизнь ефрейтора – совсем не сахар, ведь над ним столько всяческого начальства. Так и бедный мотонейрон получает сигналы со всех уровней мозга, и сигналы часто противоречивые («прыгать – нет, не прыгать»). А отвечать за все придется именно ему, стрелочнику. Это же может быть не «прыгать через скакалку», а вовсе даже «прыгать из-под колес самосвала». Не позавидуешь, не правда ли? – А теперь представьте, что у этого бедного стрелочника несколько десятков тысяч начальников! Именно так, ведь мотонейроны спинного мозга – самые конвергентные нейроны в нервной системе. Не зря великий шотландский физиолог Чарльз Шеррингтон в начале ХХ века ввел понятие общего конечного пути, которое сегодня часто «воронкой называют Шеррингтона». Откровенно говоря, задолго до него Чарльз Скотт Иван Михайлович Шеррингтон Сеченов эту концепцию высказывал наш еще великий земляк уже более Иван Михайлович Сеченов. Но суть в том, что оба настаивали, что любые события, происходящие в нашем мозгу, рано или поздно закончатся сокращением мышц, которым управляет кто? – Правильно, мотонейроны. И.М. Сеченов даже написал такую эпатажную фразу: «Мысль есть незавершенный рефлекс». Незавершенный, потому что мышцы почему-то не сократились. 3. Облегчение. Этот принцип проявляется в более мощных модулях, состоящих из десятков и сотен нейронов. Сначала договоримся о странной графике. Присутствующий в квадрате модуль состоит из 12 нейронов, расположенных в 3 колонки по 4 нейрона. Чтобы не загружать схему множеством ветвящихся аксонов, я ввел условные обозначения. Левая и центральная колонки иннервируются от нейрона А, правая и центральная – от нейрона В. То есть нейроны центральной колонки соответствуют нейрону С из раздела про конвергенцию. А вот дальше – правила работы этого модуля. Пусть от нейрона А надпорогово активируются только 3 нейрона левой колонки, от нейрона В – три нейрона правой колонки. А при их совместном синхронном воздействии добавляются еще 3 конвергентных нейрона средней колонки. А дальше – простая арифметика. ВПСПА→3ПДМ, ВПСПВ→3ПДМ, ∑ВПСПА+В→9ПДМ, то есть при раздельном последовательном воздействии на модуль 3+3=6 активированных нейронов в модуле, а при синхронном – 3+3+3=9 активированных нейронов. Следовательно, синхронная стимуляция облегчает прохождение сигнала через модуль. 4. Окклюзия. Окклюзия, вообще-то, медицинский, даже хирургический термин, означающий закупорку (окклюзия коронарных артерий ведет к инфаркту миокарда). Но в нейрональных модулях он обретает особую окраску. Обратите внимание: этот модуль по структуре (распределению синаптичеких связей) аналогичен предыдущему. А вот функционально он отличается. Дело в том, что нейроны центральной колонки надпорогово возбуждаются и при активации нейрона А, и при активации нейрона В. То есть конвергенция ничего не добавляет к активности этих нейронов. Вот дальше – парадокс. Посмотрите сами: если активируется только нейрон А в модуле возбуждаются 2 нейрона левой колонки и все 4 – центральной. При активации входа В происходит то же самое. А теперь просто посчитайте, что будет, если оба входа (А и В) активируются синхронно. Удивительно, правда? Давайте запишем: ВПСПА→6ПДМ, ВПСПВ→6ПДМ, ∑ВПСПА+В→8ПДМ, то есть при раздельном последовательном воздействии на модуль 6+6=12 активированных нейронов в модуле, а при синхронном – 2+4+2=8 активированных нейронов. Следовательно, синхронная стимуляция не только не облегчает прохождение сигнала через модуль, а напротив, ухудшает его по сравнению с раздельным последовательном воздействием нейронов А и В. 5. Латеральное торможение. Перед вами – модуль, характеризующий механизм латерального торможения. Коричневыми кружками обозначены некие рецепторы (например, клетки сетчатки глаза), от которых сигнал поступает на первую линию активирующих нейронов в модуле (зеленые кружки с зелеными линиями, обозначающими активирующие аксоны и синапсы). Красные кружочки – тормозные нейроны синапсами. Благодаря с соответствующими тому, что аксонами латеральные (то и тормозными есть «боковые», находящиеся по краям каждого ряда: самый левый и самый правый) нейроны в этой своеобразной пирамиде на каждом уровне получают большее тормозное влияние, чем возбуждающее, формируется тормозная зона вокруг центральной магистрали. В результате в трехмерной объемной нейронной сети создается своего рода изолированный канал передачи сигнала, ведь возбуждения с соседних сетей будут «вязнуть» в тормозной латеральной зоне. Это принципиально важно для мозга, ведь в толще серого вещества очень трудно создать изолированный канал передачи информации: в сером веществе с миелинизацией не все в порядке. Все эти принципы постоянно используются нейронными сетями нашего мозга, являясь простейшим инструментом построения целостной системы формирования информационных образов.