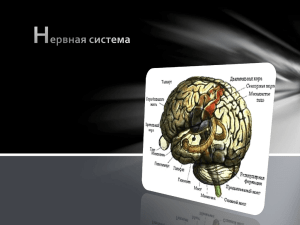
")
1. Цитология 1 Возникновение и развитие гистологии и цитологии как самостоятельных наук. Вклад отечественных и зарубежных учёных в развитие гистологии Гистология— наука о строении, развитии и жизнедеятельности тканей животных организмов. Развитие гистологии. В истории учения о тканях и микроскопическом строении органов следует различать три периода: 1-й — домикроскопический (продолжительностью около 2000 лет), 2-й — микроскопический (около 300 лет), 3-й — современный,сочетающий достижения в области электронной микроскопии, иммуноцитохимии, цитофотометрии и др. (с середины XX столетия). Первый период, наиболее продолжительный (с IV в. до н.э. и до середины XVII в.), является собственно предысторией гистологической науки, основанной на макроскопической технике. Вследствие этого в одну группу попадали иногда такие различные ткани, как нервная и соединительная (нерв и сухожилие), поэтому в середине XVII в., когда английским физиком Р. Гуком был усовершенствован микроскоп (1665), позволивший изучить тонкое строение тканей растений и животных, начинается второй период в учении о тканях. Роберт Гук ввел понятие «клетки»;Биша ввел понятия ткань и система; Гейзингер «Система гистологии»;Теодор Шванн – создал клеточные теории; Лейдиг и Келлинер создали классификацию тканей;Мари Франсуа Ксавье заложил понятие ткань, описал в своих трудах «Трактат о мембранах и оболочках» ; Третий период с 1665 по 1950 характеризуется широким и комплексным использованием многих методов исследования, и прежде всего электронной микроскопии, метода замораживания — скалывания, электронно-микроскопической цитохимии, количественных методов и др. Научно-технический прогресс, успехи развития методов исследования позволили дойти до анализа макромолекулярного уровня организации клеток и неклеточных структур, уточнить представления о процессах дифференцировки, регенерации, передаче наследственных признаков и др. Благодаря этому были созданы основы ультрамикроскопической цитологии и гистологии и разрабатываются проблемы молекулярной биологии. Вклад отечественных ученых в гистологии: 1)МЕЧНИКОВ один из основоположников эволюционной эмбриологии, создание фагоцитарной теории, иммунитета 2)КОВАЛЕВСКИЙ один из основоположников сравнительной эволюционной эмбриологии, обосновал теорию зародышевых листков 3)ЗАВАРЗИН-создатель сравнительной эволюции гистологии, теории параллелизмов в эволюции ткани 4)ХЛОПИН-создатель дивергентной теории эволюции тканей, филоонтогенетической классификации тканей. 2 Основные положения клеточной теории. Роль клеточной теории в развитие гистологии и медицины. Вклад отечественных и зарубежных ученых в учение о клетке. Клеточная теория. В настоящее время клеточная теория гласит: 1) клетка является наименьшей единицей живого, 2) клетки разных организмов принципиально сходны по своему строению, 3) размножение клеток происходит путем деления исходной клетки, 4) многоклеточные организмы представляют собой сложные ансамбли клеток и их производных, объединенные в целостные интегрированные системы тканей и органов, подчиненные и связанные между собой межклеточными, гуморальными и нервными формами регуляции. Основа клеточной теории связана с именами Матиаса Якоба Шлейдена и Теодора Шванна. Значение клеточной теории для медицины • Клетка – единица патологии (практически все болезни связаны с изменениями структуры и функции клеток) – в организме человека 200 типов клеток • Нарушение структуры и функции клеток – причины и следствие патологических процессов. ВКЛАД -Роберт Гук ввел понятие «клетки»;-1800-Биша ввел понятия ткань и система; -Гейзингер «Система гистологии»;;-Теодор Шванн – создал клеточные теории; -Лейдиг и Келлинер создали классификацию тканей;Открыли клеточный центр -Комплекс Гольджи;Мари Франсуа Ксавье 1771-1802 заложил понятие ткань, описал в своих трудах «Трактат о мембранах и оболочках»; 3. Понятие о клетке как наименьшей единице живого, являющейся основой строения эукариотических организмов. Общий план строения клеток эукариот. Неклеточные структуры организма. Основными компонентами ядра неделящейся клетки является ядерная оболочка ,хроматин,ядрышко,ядерный сок(кареоплазма) Клетка - структурно-функциональная единица живого. Для всех эукариотических клеток характерно наличие следующих структур: 1) Клеточная мембрана - это органоид, ограничивающий содержимое клетки от окружающей среды, также регулирует транспорт веществ из клетки и в неё. 2) Цитоплазма - внутренняя среда клетки, в которую погружены все органоиды клетки и различные продукты жизнедеятельности. 3) Эндоплазматическая сеть (ЭПС) - система трубочек и расширений, которая связана в единое целое с ядерной мембраной и клеточной мембраной. Данный органоид обеспечивает транспорт веществ внутри клетки, а также осуществляет синтез различных веществ, например, белков, который принадлежит шероховатой ЭПС, а также синтез жиров и углеводов - гладкая ЭПС. 4) Рибосомы - органоиды округлой или грибовидной формы, состоящие из двух субъединиц, в них синтезируются белки. 5) Митохондрии - двумембранные органоиды клетки, участвующие в энергетическом процессе. Их ещё иначе называют энергетическими станциями клетки. 6) Аппарат Гольджи органоиды, состоящие из стопки плоских цистерн, которые участвуют во всех обменных процессах в клетке, а также в образовании лизосом. 7) Лизосомы - одномембранные органоиды клетки, выполняющие множество функций, связанных с распадом клеточных структур или молекул. 8) Клеточный центр - немембранный органоид, состоящий из двух центриолей, который принимает участие в делении клеток животных и низших растений. 9) Ядро - органоид, окруженный двойной мембраной, которая пронизана ядерными порами. Содержит также структуру, называемую ядрышком. Ядро хранит в себе наследственную информацию Среди неклеточных структур различают ядерные, содержащие ядра и возникают путем слияния клеток или вследствие незавершенного разделения их, и безъядерные - продукт деятельности определенных видов клеток. К ядерным неклеточных структур относятся симпласты и синцитий. Симпластам - неклеточная структура, которая является массой нерасчлененной на клетки цитоплазмы с большим количеством ядер. Симпластичну строение имеют скелетные мышечные волокна, а также внешний слой зародышевой части плаценты. Синцитий, или суклиття (клеточная сетка, сетчатый симпластам) - это группа клеток, которые объединены в единое целое цитоплазматическими мостиками. Такая временная структура возникает при развитии мужских и женских половых клеток, когда разделение клеточного тела не заканчивается.К безъядерных неклеточных структур относятся волокна и основное (аморфное) вещество соединительной ткани, продуцируемых одним из типов клеток - фибробластами. Аналогами основного вещества есть такие жидкие среды, как плазма крови и жидкая часть лимфы. 4. Биологическая мембрана. Структурно-функциональная характеристика, основные свойства и функции. Понятие о компартментализации клетки и её функциональное значение. В липидном бислое фосфолипидные гидрофобные группы обращены во внутрь, а гидрофильные наружу. Белковые молекулы (интегральные белки) вмонтированы в плазмалемму. Если белковая молекула пронзает всю толщу мембраны – это трансмембранный белок. Если белковая молекула прикрепляется к поверхности мембраны – это периферические белки (внутренние – белки цитоскелета, наружные – рецепторные белки). Трансмембранные белки образуют ионные каналы. Мембранные белки: прикрепляют филаменты цитоскелета к клеточной мембране; прикрепляют клетки к экстрацеллюлярному матриксу (адгезионные молекулы); транспортируют молекулы в клетку или из неё (белкипереносчики, белки мембранных насосов, белки ионных каналов); действуют как рецепторы химического взаимодействия между клетками; обладают специфической ферментативной активностью. В клеточной мембране также присутствуют гликолипиды, холестерин (ограничивает латеральную текучесть фосфолипидов, делает мембрану менее текучей и более стабильной). Гликолипиды вовлечены в межклеточные взаимодействия. На поверхности выступают и молекулы углеводов, соединённые либо с гликолипидами, либо с белками. Между хвостами противолежащих молекул фосфолипидов есть лишь слабые гидрофобные связи, удерживающие две половины мембраны вместе. При замораживании-скалывании клеточная мембрана расщепляется вдоль так, что большая часть интегральных белков отходит к внутреннему листку, и лишь некоторые из них - наружному. Функции: установление структурной целостности клетки; селективная проницаемость; регуляция межклеточных взаимодействий; узнавание, через рецепторы, антигенов, повреждённых клеток, чужих клеток; трансдукция внешнего химического и физического сигнала во внутриклеточное событие; служит разделом сред между цитоплазмой и внешним окружением; образует транспортные системы для особых молекул, как, например, глюкоза. Гликокаликс – тонкая филаментозная сеть на поверхности клеток, отходящая от наружного листка плазмалеммы, состоящая из олигосахаридов, ковалентно связанных с гликолипидами и гликопротеинами плазмалеммы. Играет важную роль в определении иммунологических свойств клетки и её взаимодействии с другими клетками. 5. Биологическая мембрана. Структурно-функциональная характеристика, основные свойства и функции. Структурно-функциональная характеристика различных видов межклеточных соединений. Цитолемма(плазмолемма) выполняет разграничительную функцию и регулирует движение ионов и молекул в клетку и из клетки, а также участвует в процессах фагоцитоза, пиноцитоза и экзоцитоза. Цитолемма представляет собой элементарную биологическую мембрану, состоит из двойного слоя липидов и белков интегральных, полуинтегральных и периферических (транспортных, или белков-переносчиков). Кроме того, с липидами и белками связаны молекулы углеводов, образуя с ними сложные соединения - гликолипиды и гликопротеиды. Они формируют надмембранный комплекс - гликокаликс, в составе которого есть структуры, способные специфически связывать определенные химические вещества и называемые рецепторами. С внутренней стороны мембраны располагается подмембранный (субмембранный) комплекс, включающий в себя микрофиламенты. микрофиб-риллы и микротрубочки цитоскелета, а также актомиозиновый комплекс. Специализированными структурами цитолеммы являются различные типы межклеточных соединений, а также выросты цитоплазмы -микроворсинки или более сложные по строению реснички и жгутики. Различают следующие типы межклеточных соединений: простые межклеточные соединения (зубчатые и пальцевидные); пятна сцепления, или десмосомы; плотные соединения; пояски сцепления, или лентовидные десмосомы; щелевидные соединения, или нексусы, в области которых из клетки в клетку могут проникать небольшие молекулы и ионы, но не белки. По нексусам может передаваться возбуждение в сердечной или гладкой мышечной ткани. Синаптические соединения, или синапсы, характерны для нервной ткани, обычно они осуществляют одностороннюю передачу возбуждения или торможения между клетками. Образование новых мембран в клетке идет с участием гладкой (липидная часть) и гранулярной (белковые компоненты) эндоплазматической сети. Это наблюдается после деления клетки, при повреждении (точнее, при восстановительных процессах после повреждения), при гипертрофии клетки. 6. Специализированные структуры клеточной поверхности: микроворсинки, реснички, жгутики, базальные инвагинации. Их строение и функции. Микроворсинки - структуры клетки, располагающиеся на ее свободной поверхности и выступающие во внеклеточное пространство. Эти выпячивания значительно расширяют поверхность клетки. Микроворсинки активно всасывают и (или) активно переваривают вещества на поверхности клеток. Микроворсинкам присущи некоторым популяциям эпителиоцитов. Нередко для клеток, имеющих много микроворсинок, характерна высокая ферментативная активность и счет белков, прикрепляющихся к гликокаликсу (столбчатые энтероциты кишечника, тироциты щитовидной железы и т. д.). Микроворсинки сформированы клеточной мембраной к гиалоплазмой. Реснички и жгутики. Реснички и жгутики на ультраструктурном уровне имеют сходные принципы строения, ко могут иметь разные функции. Реснички перемещают поверхностный субстрат полого органа, тогда как жгутик сперматозоида позволяет передвигаться самой клетке. Реснички состоят из базалъного тельца и выступающей из клетки аксонемы. Базальное тельце по строению аналогично дочерней центриоле и состоит из девяти триплетов микротрубочек. Базальные инвагинации (впячивания). Базальные впячивания – это внедрения цитолеммы в цитоплазму, прилежащие к базальной мембране или иной плотной структуре, например, впячивание цитолеммы на остеокласте со стороны разрушаемой (резорбцируемой) кости, существенно расширяющие поверхность клетки с внутренней (базальной) поверхности. Нередко в зонах впячиваний цитолеммы видны многочисленные мнтохондрии. Совокупность базальных инвагинаций и митохондрий формирует базальную исчерченность. Наличие базальной исчерченности признак активных процессов трансмембранного перекоса веществ, когда наряду с увеличением поверхности значение имеет и высокий уровень энергетического потребления, обеспечиваемый АТФ. 7. Органеллы цитоплазмы: понятие и классификация. Строение органелл, участвующих в биосинтезе веществ в клетках. Органеллы – постоянные структурные элементы цитоплазмы клетки, имеющие специфическое строение и выполняющие определенные функции. Классификация: 1)общие органеллы, присущие всем клеткам делятся на: а)мембранные органеллы: митохондрии, эндоплазматическая сеть, пластинчатый комплекс, лизосомы, пероксисомы; б)немембранные органеллы: рибосомы, клеточный центр, микротрубочки, микрофибриллы, микрофиламенты.2) Специальные органеллы, имеющиеся в цитоплазме только определенных клеток, делятся на: а)цитоплазматические миофибриллы, нейрофибриллы, тонофибриллы; б)органеллы клеточной поверхности – реснички, жгутики. Органеллы общего значения мембранного строения: Эндотазматическая сеть (ЭПС) — система канальцев и уплощенных цистерн. Гранулярная – с рибосомами она способна к синтезу белка. Гладкая связана с метаболизмом углеводов и липидов. Здесь же возможен син­тез стероидных гормонов, обезвреживание снотворных веществ и канцерогенов, депонирование ионов кальция (в мышечной ткани). ЭПС является системой внутриклеточного транспорта. Комплекс Гольджи уплощенные цистерны, а также микропузырьки, крупные вакуоли и секреторные гранулы. Функции орга­ноида- упакФовка, конденсация и выведение белковых секретов, участие в синтезе углеводов, гликопротеинов и гликозаминогликанов, образование первичных лизосом, участие в эндо- и экзоцитозе. Лизосамы - органеллы с защитной и пищеварительной функцией, способные расщеплять своими фермента­ми вещества и структуры как эндогенного, так и экзогенного происхождения. Пероксисомы напоминают лизосомы, но вместо гидролаз содержат оксидазы и каталазу, способную разру­шать токсичную для клеток перекись водорода. Митохондрии - имеющие двухмембранное строение. Внутренняя мембрана образует кристы, увеличивающие ее площадь, полость внутренней камеры заполнена митохондриаль-ным матриксом, в котором содержатся АТФ-сомы (комплексы ферментов фосфорилирования). Функции митохондрий: обеспечение клетки энергией, синтез АТФ, молекулы которого депонируются в мито­хондриях и тратятся по мере надобности; внутриклеточное дыхания; автономный синтез белка. Органеллы общего значения немембранного строения: Рибосамы - гранулы рибонуклеопротеидов, в состав которых входят белки и молекулы РНК: состоит из большой и малой субъединиц. Функция рибосом - синтез белка. Клеточный центр (центросома) — две центриоли, окруженные центросферой (светлой зоной), от которой радиально отходят тонкие фибриллы, формирующие астросферу. Центриоли - два цилиндра, лежащие под пря­мым углом друг к другу. Стенка цилиндра состоит из девяти триплетов микротрубочек. Центриоли служат центрами формирования всех мик­ротрубочек клетки. Микротрубочки построены из белка тубулина. 8. Органеллы цитоплазмы: понятие и классификация. Строение органелл, участвующих во внутриклеточном пищеварении, защитных и обезвреживающих реакциях. Органеллы – постоянные структурные элементы цитоплазмы клетки, имеющие специфическое строение и выполняющие определенные функции. Классификация: 1)общие органеллы, присущие всем клеткам делятся на: а)мембранные органеллы: митохондрии, эндоплазматическая сеть, пластинчатый комплекс, лизосомы, пероксисомы; б)немембранные органеллы: рибосомы, клеточный центр, микротрубочки, микрофибриллы, микрофиламенты.2) Специальные органеллы, имеющиеся в цитоплазме только определенных клеток, делятся на: а)цитоплазматические миофибриллы, нейрофибриллы, тонофибриллы; б)органеллы клеточной поверхности – реснички, жгутики. Лизосомы. Представляют собой небольшие округлые тельца. От Цитоплазмы каждая лизосома отграничена мембраной. Внутри лизосомы находятся ферменты, расщепляющие белки, жиры, углеводы, нуклеиновые кислоты. К пищевой частице, поступившей в цитоплазму, подходят лизосомы, сливаются с ней, и образуется одна пищеварительная вакуоль, внутри которой находится пищевая частица, окруженная ферментами лизосом. Вещества, образовавшиеся в результате переваривания пищевой частицы, поступают в цитоплазму и используются клеткой. Обладая способностью к активному перевариванию пищевых веществ, лизосомы участвуют в удалении отмирающих в процессе жизнедеятельности частей клеток, целых клеток и органов. Образование новых лизосом происходит в клетке постоянно. Ферменты, содержащиеся в лизосомах, как и всякие другие белки синтезируются на рибосомах цитоплазмы. Затем эти ферменты поступают по каналам эндоплазматической сети к аппарату Гольджи, в полостях которого формируются лизосомы. В таком виде лизосомы поступают в цитоплазму. Пероксисомы — это органеллы, напоминающие лизосомы, но содержащие ферменты, необходимые для синтеза и разрушения эндогенных перекисей — пероксидазу, каталазу и другие, всего до 15. В Электронном микроскопе представляют сферические или эллипсоидные пузырьки с умеренно плотной сердцевиной. Образуются пероксисомы путем отделения пузырьков от гладкой ЭПС. В эти пузырьки затем мигрируют ферменты, которые синтезируются отдельно в цитозоле или в гранулярной ЭПС. Функции пероксисом: 1. Являются, наряду с митохондриями, органеллами утилизации кислорода, В результате в них образуется сильный окислитель Н2О2. 2. Расщепление при помощи фермента каталазы избытка перекисей и, таким образом, защита клеток от гибели. 3. Расщепление при помощи синтезируемых в самих пероксисомах перекисей токсических продуктов, имеющих экзогенное происхождение (детоксикация). Такую функцию выполняют, например, пероксисомы печеночных клеток, клеток ночек. 4. Участие в метаболизме клетки: ферменты пероксисом катализируют расщепление жирных кислот, участвуют в обмене аминокислот и других веществ. 9. Органеллы цитоплазмы: понятие и классификация. Строение органелл, участвующих в процессах выведения веществ из клетки. Органеллы – постоянные структурные элементы цитоплазмы клетки, имеющие специфическое строение и выполняющие определенные функции. Классификация: 1)общие органеллы, присущие всем клеткам делятся на: а)мембранные органеллы: митохондрии, эндоплазматическая сеть, пластинчатый комплекс, лизосомы, пероксисомы; б)немембранные органеллы: рибосомы, клеточный центр, микротрубочки, микрофибриллы, микрофиламенты.2) Специальные органеллы, имеющиеся в цитоплазме только определенных клеток, делятся на: а)цитоплазматические миофибриллы, нейрофибриллы, тонофибриллы; б)органеллы клеточной поверхности – реснички, жгутики. Включения цитоплазмы — необязательные компоненты клетки, возникающие и исчезающие в зависимости от метаболического состояния клеток. Различают включения трофические, секреторные, экскреторные и пигментные. К трофическим включениям относятся капельки нейтральных жиров, которые могут накапливаться в гиалоплазме. В случае недостатка субстратов для жизнедеятельности клетки эти капельки могут резервироваться. Другим видом включений резервного характера является гликоген — полисахарид, откладывающийся также в гиалоплазме. Отложение запасных белковых гранул обычно связано с активностью эндоплазматической сети. Так, запасы белка вителлина в яйцеклетках амфибии накапливаются в вакуолях эндоплазматической сети. Секреторные включения — обычно округлые образования различных размеров, содержащие биологически активные вещества, образующиеся в клетках в процессе синтетической деятельности. Экскреторные включения не содержат каких-либо ферментов или других активных веществ. Обычно это продукты метаболизма, подлежащие удалению из клетки. Пигментные включения могут быть экзогенными (каротин, пылевые частицы, красители и др.) и эндогенными (гемоглобин, гемосидерин, билирубин, меланин, липофусцин). Наличие их в цитоплазме может изменять цвет ткани, органа временно или постоянно. Нередко пигментация ткани служит диагностическим признаком. 10. Органеллы цитоплазмы: понятие и классификация. Строение органелл, участвующих в энергопроизводстве. Органеллы – постоянные структурные элементы цитоплазмы клетки, имеющие специфическое строение и выполняющие определенные функции. Классификация: 1)общие органеллы, присущие всем клеткам делятся на: а)мембранные органеллы: митохондрии, эндоплазматическая сеть, пластинчатый комплекс, лизосомы, пероксисомы; б)немембранные органеллы: рибосомы, клеточный центр, микротрубочки, микрофибриллы, микрофиламенты.2) Специальные органеллы, имеющиеся в цитоплазме только определенных клеток, делятся на: а)цитоплазматические миофибриллы, нейрофибриллы, тонофибриллы; б)органеллы клеточной поверхности – реснички, жгутики. В цитоплазме протекают основные процессы обмена веществ, она объединяет в одно целое ядро и все органоиды, обеспечивает их взаимодействие, деятельность клетки как единой целостной живой системы. К ним относятся митохондрии. Они представляют собой полуавтономные органеллы и аппарат синтеза АТФ за счет энергии, получаемой при окислении органических соединений. Эти органеллы способны перемещаться по цитоплазме, сливаться одна с другой, делиться. Форма и размеры различны, число их зависит от активности клетки. Чаще всего это тельца длиной 1-10 мкм, толщиной 0,5 мкм. Митохондрии состоят из наружной и внутренней мембран, разделенных межмембранным пространством, и содержат митохондриальный матрикс, в который обращены складки внутренней мембраны (кристы). Наружная митохондриальная мембрана напоминает плазмолемму, содержит много молекул специализированных транспортных белков (например, порин), формирующих каналы, обеспечивающие высокую проницаемость. На ней находятся рецепторы, распознающие белки, которые переносятся через обе митохондриальные мембраны в зонах их слипания. Функции митохондрий 1. Обеспечение клетки энергией в виде АТФ. 2. Участие в биосинтезе стероидных гормонов (некоторые звенья биосинтеза этих гормонов протекают в митохондриях). В таких клетках – митохондрии со сложными крупными трубчатыми кристами. 3. Депонирование кальция. 4. Участие в синтезе нуклеиновых кислот. Продолжительность существования митохондрий – около 10 суток. Их разрушение происходит путем аутофагии. Образование новых митохондрий происходит путем перешнуровки предшествующих. 11. Строение органелл специального назначения (микроворсинки, базальные складки, реснички, жгутики, тонофибриллы, миофибриллы и нейрофибриллы). Органеллы спец. назначения– это постоянно присутствующие и обязательныедля отдельных клеток микроструктуры, выполняющие особые функции, которыеобеспечивают специализацию ткани и органа. К ним относят: реснички, жгутики, микроворсинки, миофибриллы. Реснички и жгутики– это специальные органеллы движения, встречающиеся в некоторых клетках различных организмов. Ресничка представляет собой цилиндрический вырост цитоплазмы. Внутри выроста располагается аксонема ( осевая нить) , проксимальная часть реснички( базальное тело) погружена в цитоплазму. Систему микротрубочек реснички описывают по формуле – (9х2) + 2. Основной белок реснички- тубулин. Тонофибриллы- тонкие белковые волокна, обеспечивающие сохранность формы в некоторых эпителиальных клетках. Тонофибриллы обеспечивают механическую прочность клеток. Миофибриллы— это органеллы клетокпоперечнополосатых мышц, обеспечивающие их сокращение. Служат для сокращениймышечных волокон. Миофибрилла — это нитевидная структура, состоящая изсаркомеров. Каждый саркомер имеет длину около 2 мкм и содержит два типа белковых филаментов: тонкиемикрофиламентыизактинаи толстые филаменты измиозина. Границы между филаментами (Z-диски) состоят из особых белков, к которым крепятся ±концы актиновых филаментов. Миозиновые филаменты также крепятся к границам саркомера с помощью нитей из белкатитина(тайтина). С актиновыми филаментами связаны вспомогательные белки —небулини белкитропонинтропомиозиновогокомплекса. У человека толщина миофибрилл составляет 1-2 мкм, а их длина может достигать длины всей клетки (до нескольких сантиметров). Одна клетка содержит обычно несколько десятков миофибрилл, на их долю приходится до 2/3 сухой массы мышечных клеток. 12. Включения цитоплазмы. Гиалоплазма. Физико-химические свойства, химический состав. Участие включений цитоплазмы в клеточном метаболизме. Включения цитоплазмы — необязательные компоненты клетки, возникающие и исчезающие в зависимости от метаболического состояния клеток. Гиалоплазма (матрикс) заполняет пространство между плазмалеммой, ядерной оболочкой и другими внутриклеточными структурами. Это тонкозернистое, полупрозрачное, вязкое, студенистое вещество цитоплазмы. Химический состав. Гиалоплазма – это коллоидный раствор с высоким содержанием воды и белков. Гиалоплазма способна переходить из золеобразного (жидкого) состояния в гелеобразное. Состав гиалоплазмы определяет осмотические свойства клетки. Н2О 70 – 75%,белки 10 – 20%,липиды 1 – 5%,углеводы 0,2 – 2%,нуклеиновые кислоты 1 – 2%,минеральные соединения 1 – 1,5%,АТФ и другие низкомолекулярные органические вещества 0,1 – 0,5%. Физико-химические свойства гиалоплазмы обусловлены ее коллоидным характером. Они определяются наличием в ней множества частиц, в совокупности образующих огромную поверхность взаимодействия со средой, что обеспечивает прохождение разнообразных физико-химических процессов. Благодаря силе поверхностного натяжения, возникающей на микроскопическом комочке гиалоплазмы, осуществляется процесс адсорбции— концентрации одного вещества на поверхности другого. В зависимости от увеличения, даваемого микроскопом, гиалоплазма представляется гомогенной или зернистой, гранулированной. Размер гранул близок к размеру макромолекул. 13. Ядро клетки. Характеристика ядра как генетического центра клетки. Роль ядра в хранении и передаче генетической информации и в синтезе белка. Ядро клетки — структура, обеспечивающая генетическую детерминацию и регуляцию белкового синтеза.23 пары хромосом, ядра соматической клетки содержат ядерный геном. Реализация генетической информации (транскрипция, процессинг, трансляция, посттрансляционная модификация) и другие функции ядра происходят при участии ДНК и разных видов РНК. Структурные компоненты ядра: хроматин, ядерная оболочка, ядрышко, нуклеоплазма. ХРОМАТИН Термином «хроматин» обозначают комплекс ядерной ДНК с белками (гистоны, негистоновые). Различают гетеро- и эухроматин. Гетерохроматин. Гетерохроматин — транскрипционно неактивный, конденсированный хроматин интерфазного ядра. В СМ — базофильные глыбки. в ЭМ —скопления плотных гранул Располагается преимущественно по периферии ядра и вокруг ядрышек. Типичный пример гетерохроматина — тельце Барра. Эухроматин. Эухроматин — транскрипционно активная и менее конденсированная часть хроматина, локализуется в более светлых участках ядра между гетерохроматином. ЯДЕРНАЯ ОБОЛОЧКА В состав ядерной оболочки входят наружная и внутренняя ядерные мембраны перинуклеарные цистерны ядерная пластинка, ядерные поры. На поверхности наружной ядерной мембраны расположены рибосомы, где синтезируются белки, поступающие в перинуклеарные цистерны, рассматриваемые как часть гранулярной эндоплазматической сети. Внутренняя ядерная мембрана отделена от содержимого ядра ядерной пластинкой. Ядерная пластинка толщиной 80—300 нм участвует в организации ядерной оболочки перинуклеарного хроматина, может разделять комплексы ядерных пор и дезинтегрировать ядро в ходе митоза; содержит белки промежуточных филаментов — ламины А, В и С. Ядерная пора имеет диаметр 80—150 нм, содержит канал поры и комплекс ядерной поры. Содержимое ядра сообщается с цитозолем через 3—4 тысячи специализированных коммуникаций — ядерных пор, осуществляющих диффузию воды, ионов и транспорт множества макромолекул (в т.ч. молекул РНК) между ядром и цитоплазмой. ЯДРЫШКО Ядрышко — компактная структура в ядре интерфазных клеток. В ядрышке различают фибриллярный центр слабоокрашенный компонент (ДНК, кодирующая рРНК), фибриллярный компонент,где протекают ранние стадии образования предшественников рРНК, состоит из тонких (5 нм) рибонуклеопротеиновых фибрилл и транскрипционно активных участков ДНК. НУКЛЕОПЛАЗМАЯдерный матрикс содержит сеть рибонуклеопротеинов, ядерные рецепторы, ферменты (АТФаза, ГТФаза, НАД-пирофосфатаза, ДНК- и РНК-полимеразы) и множество других молекул, часто образующих ассоциации — ядерные частицы. В матриксе происходят транскрипция и процессинг мРНК и рРНК. Ядерные рецепторы, онкогены, факторы транскрипции, белки теплового шока, вирусные ДНК влияют на транскрипцию и процессинг РНК. 14. Хроматин. Строение и химический состав. Хроматин как форма существования хромосом в неделящемся ядре. Хроматин представляет собой вещество, хорошо восприни мающее краситель (хромос), откуда и произошло его название. Хроматин состоит из хроматиновых фибрилл толщиной20—25км, которые могут располагаться в ядре рыхло или компактно. На этом основании можно выделить эухроматин — рыхлый (или деконденсированный) хроматин, слабо окрашиваемый ос новными красителями, и гетерохроматин — компактный (или конденсированный) хроматин, хорошо окрашиваемый основны ми красителями. При подготовке клетки к делению в ядре происходит спира лизация хроматиновых фибрилл и превращение хроматина в хро мосомы. После деления в ядрах дочерних клеток происходит деспирализация хроматиновых фибрилл, и хромосомы снова пре образуются в хроматин. Таким образом, хроматин и хромосомы являются различными состояниями одного и того же вещества. По химическому строению хроматин состоит из: 1)дезоксирибонуклеиновой кислоты (ДНК) — 40%;2)белков — около 60%;3)рибонуклеиновой кислоты (РНК) — 1%. Ядерные белки представлены двумя формами:1)щелочными (гистоновыми) белками — 80—85%; 2)кислыми белками — 15—20%. Хроматин – это деспирализованная форма существования хромосом в неделящемся ядре. Его химическую основу составляет дезоксирибонуклеопротеин – комплекс ДНК с гистоновыми и негистоновыми белками. При этом до момента репликации ДНК каждая хромосома содержит лишь одну линейную молекулу ДНК. Хроматин соответствует хромосомам, которые в интерфазном ядре представлены длинными перекрученными нитями и неразличимы как индивидуальные структуры. Выраженность спирализации каждой из хромосом неодинакова по их длине. Реализацию генетической информации осуществляют деспирализованные участки хромосом. 15. Морфология митотических хромосом. Кариотип. Кариоти́п — совокупность признаков (число, размеры, форма и т. д.) полного набора хромосом, присущая клеткам данного биологического вида (видовой кариотип), данного организма (индивидуальный кариотип) или линии (клона) клеток. Понятие К. введено сов. генетиком Г. А. Левитским (1924). К. — одна из важнейших генетических характеристик вида, т.к. каждый вид имеет свой К., отличающийся от К. близких видов (на этом основана новая отрасль систематики — так называемая кариосистематика).Постоянство К. в клетках одного организма обеспечивается митозом, а в пределах вида — мейозом. К. организма может изменяться, если половые клетки (гаметы) претерпевают изменения под влиянием мутаций. Хромосома делящейся клетки состоит из 2 хроматид,соединенных между собой перетяжкой. Последняя- неспирализованный участок ДНК. Снаружи хромосомы покрыты белковой оболочкой из гистонов. Хромосомы состоят из ДНК(90%) и РНК(10%). Набор хромосом бывает диплоидный(2n2c) – у соматических клеток и гаплоидный(1n1c)- у половых клеток. Функция хромосом – синтез нуклеиновых кислот. Боковые петли хромосом в вытянутом состоянии могут достигать 30 мкм. При их компактизации (спирализации) образуются структуры промежуточного характера — так называемые хромонемные фибриллы. 16. Ядрышко. Ядерная оболочка. Кариоплазма. Я́дрышко — немембранный внутриядерный субкомпартмент, присущий всем без исключения эукариотическим организмам. Представляет собой комплекс белков и рибонуклеопротеидов, формирующийся вокруг участков ДНК, которые содержат гены рРНК— ядрышковых организаторов. Основная функция ядрышка — образование рибосомных субъединиц. Ядерная оболочка - мембранный барьер, отделяющий ядро от цитоплазмы. Она контролирует перемещение макромолекул между нуклеоплазмой и цитозолем, участвует в заякоревании хромосом и цитоскелета, являясь частью регуляторного механизма экспрессии у эукариот. В состав ядерной оболочки входят мембрана, ядерный поровый комплекс и ламина. Ядерная оболочка образована внешней и внутренней мембранами. Наружная мембрана переходит в шероховатый эндоплазматический ретикулум, и обеспечивает присоединение структурных элементов цитоплазмы. Вутренняя выстлана белками – ламининами, образующими ядерную пластинку, которая закрепляет различные ядерные структуры. Между мембранами располагается перинуклеарное пространство. Кариоплазма или нуклеоплазма. Это микроскопически бесструктурное вещество ядра. Содержит различные белки, нуклеопротеины, гликопротеины, ферменты и соединения, участвующие в процессах синтеза нуклеиновых кислот, белков и всех других веществ, входящих в состав кариоплазмы. Кариоплазма заполняет все внутреннее пространство ядра между хромосомами и ядрышком. Благодаря ей ядро обладает тургором. По долевому вкладу больше всего в кариоплазме РНК, которая относится к неядрышковым продуктам транскрипции. Это перихроматиновые фибриллы, перихроматиновые гранулы и интерхроматинвоые гранулы. Кариоплазма(ядерный сок, нуклеоплазма) в виде неструктурированной массы окружает хромосомы и ядрышки. Вязкость ядерного сока примерно такая же, как вязкость основного вещества цитоплазмы. Кислотность ядерного сока, определенная путем микроинъекции индикаторов в ядро, оказалась несколько выше, чем у цитоплазмы. Кроме того, в ядерном соке содержатся ферменты, участвующие в синтезе нуклеиновых кислот в ядре, и рибосомы. Ядерный сок не окрашивается основными красителями, поэтому его называют ахроматиновым веществом, или кариолимфой, в отличие от участков, способных окрашиваться, - хроматина. 17. Взаимодействие структур ядра и цитоплазмы в процессе синтеза белка и небелковых веществ в клетках. Структурные компоненты клетки взаимосвязаны и взаимодействуют между собой в процессе жизнедеятельности клетки. Например, при биосинтезе белка на экспорт (секреторные белки) через цитолемму внутрь клетки поступают необходимые исходные вещества (аминокислоты). В ядре в результате транскрипции образуется информационная РНК, которая поступает в цитоплазму и несёт информацию о строении будущего белка, сюда же из ядрышек доставляются субъединицы рибосом и транспортные РНК. На рибосомах гранулярной цитоплазматической сети происходит биосинтез белка и образующиеся его молекулы поступают внутрь цистерн и каналов этой сети, где образуется их вторичная и третичная структура. Затем белки транспортируются в комплекс Гольджи. Там происходит дозревание (связывание белков с углеводами и липидами), накопление и упаковка секрета в мембраны, образование крупных вакуолей и гранул секрета. Затем секреторные вакуоли и гранулы выделяются через цитолемму апикальной части клетки путём экзоцитоза. Энергию, необходимую для синтетических процессов, поставляют митохондрии. Микротрубочки и микрофиламенты цитоскелета обеспечивают перемещение в цитоплазме органелл и транспорт веществ. Изнашиваемые в ходе этих процессов органоиды разрушаются лизосомами, а вместо них образуются новые. Таким образом, большинство структурных компонентов клетки принимают участие и взаимодействуют между собой в процессе биосинтеза белка. При этом клетка функционирует как единое целое. В синтезе небелковых веществ (углеводы, липиды) также участвуют ДНК ядра, информационная РНК, свободные рибосомы, на которых образуются ферменты биосинтеза небелковых веществ. Эти ферменты поступают в гладкую эндоплазматическую сеть, где участвуют в синтезе углеводов и липидов. Эти вещества затем направляются в комплекс Гольджи, где упаковываются в секреторные гранулы, а затем выводятся наружу путём экзоцитоза. 18. Понятие о секреторном цикле в клетке. Способы выделения секреторных продуктов из клетки. Секреторным циклом называются периодические изменения железистой клетки, связанные с образованием, накоплением и восстановлением ее для дальнейшей секреции. Условно секреторный цикл разделяется на следующие фазы: I фаза поступления веществ из крови и лимфы со стороны базальной поверхности различных неорганических соединений, низкомолекулярных органических веществ (аминокислот, моносахаридов, жирных кислот и др.) и воды. II фаза секреции. В эндоплазматической сети, в зависимости от состава секрета (гранулярной или агранулярной), синтезируются секреты, которые затем перемещаются в аппарат Гольджи, где постепенно накапливаются, подвергаются химической перестройке, оформляются в виде гранул. Значительная роль в перемещении секреторных продуктов в гландулоцитах принадлежит микротрубочкам и микрофиламентам – элементам цитоскелета. Синтез секрета протекает непрерывно, однако он может накапливаться и выводится из клетки с разной интенсивностью или диффузно, или в виде гранул. III фаза выделения секрета. По способу высвобождения секрета (экструзии) железы подразделяются на железы с тремя типами секреции: мерокриновый тип (эккриновый), апокриновый и голокриновый. При мерокриновом типе секреции экструзия происходит без разрушения мембран, клетки сохраняют свою целостность (например, в слюнных железах млекопитающих). При апокриновом типе секреции апикальная часть секреторных клеток может частично разрушаться, т.е. вместе с секреторными продуктами отделяются либо апикальная часть цитоплазмы или верхушки микроворсинок (например, в клетках молочной железы). Голокриновый тип секреции сопровождается накоплением секрета в цитоплазме гландулоцита и последующим полным разрушением клеток (например, в сальных железах кожи). В наружном ростковом слое сальных желез находятся недифференцированные клетки, способные к пролиферации. IV фаза восстановления исходных размеров и структуры железистых клеток. В некоторых железах, помимо секреторных, встречаются эпителиальные клетки, обладающие способностью сокращаться – миоэпителиальные клетки, имеющие отросчатую форму. Эти клетки охватывают концевые отделы желез. При сокращении миоэпителиальных клеток сдавливаются концевые отделы желез и выводится секрет. 19. Митоз, как основной способ репродукции клеток. Репродукция клеток осуществляется путём митоза (для соматических клеток) или мейоза (при созревании половых клеток). Митоз, кариокинез или непрямое деление – универсальный способ деления любых животных клеток. При этом удвоившиеся и конденсированные хромосомы переходят в компактную форму митотических хромосом. Затем образуется веретено деления, которое обеспечивает разделение и расхождение хромосом к противоположным полюсам клетки. Митоз заканчивается делением тела клетки (цитотомия). Биологическая сущность митоза заключается в равномерном распределении генетического материала между дочерними клетками. Процесс митоза подразделяется на несколько основных фаз: профаза, метафаза, анафаза и телофаза. Профаза. ДНК в результате суперспирализации начинает выявляться под микроскопом в ядре клетки в виде палочковидных телец – хромосом. Метафаза. Заканчивается образование веретена деления, а хромосомы выстраиваются в экваториальной плоскости клетки, образуя метафазную пластинку хромосом (вид сбоку) или материнскую звезду (вид с полю сов клетки). К концу метафазы завершается процесс разделения сестринских хроматид и они остаются связанными между собой только в области центромера. Анафаза. Хромосомы растягиваются к полюсам клетки с помощью микротрубочек веретена деления со скоростью 0,2-0,5 мкм/мин, что связано с деполимеризацией и укорачиванием микротрубочек и работой белков-транслокаторов. Телофаза. Начинается с остановки разошедшихся к полюсам хромосом (ранняя телофаза) и заканчивается созданием новых интерфазных ядер и разделением материнской клетки на две дочерние в результате цитотомии (поздняя телофаза). При этом хромосомы деконденсируются, образуются ядерные оболочки и формируются новые ядрышки. 20. Мейоз. Его особенности и биологическое значение. Мейоз – способ деления, в результате которого образуются клетки с гаплоидным набором хромосом (половые клетки). Оба деления мейоза происходят как обычный митоз, однако в профазе первого деления происходит обмен генами между гомологичными хромосомами (кроссинговер), между первым и вторым делением нет интерфазы и поэтому не происходит редупликации ДНК. Биологическое значение мейоза заключается в том, что образовавшиеся мужские и женские половые клетки несут генетическую информацию от отца и матери и при слиянии этих клеток образуется зигота с диплоидным набором хромосом, несущая равное количество генетической информации от обоих родителей. 21. Способы репродукции клеток. Эндорепродукция. Основные формы, биологическое значение. Понятие о плоидности клеток. Полиплоидия. Способы репродукции клеток Митоз – универсальный способ деления соматических клеток Результат: увеличение количества идентичных клеток Амитоз – прямое деление (иногда в соматических клетках) Результат: образование двуядерных (многоядерных) клеток или дочерних генетически неполноценных клеток Мейоз – деление клетокпредшественников гамет Результат: образование гаплоидных, генетически разнородных половых клеток. Митоз деление соматических клеток Результат: увеличение количества генетически идентичных клеток; генетический материал между дочерними клетками распределяется поровну . Эндорепродукция — это последовательное многократное удвоение ДНК, в результате чего увеличивается набор хромосом, при этом хромосомы связаны тонкими нитями. Эти структуры называются политенами, характерными для клеток плаценты. Эндорепродукция - явление, связанное не с увеличением количества клеток, а с увеличением (репродукцией) генетического материала в клетке. Различают два вида эндорепродукции: эндомитоз и политению. Эндомитоз происходит при нарушении нормального хода митоза (сохранение ядерной оболочки в профазе, разрушение митотического аппарата в начале анафазы) и приводит к увеличению плоидности клетки, кратное " n ". Если в эндомитоз вступила клетка, содержавшая 2n, то образуется клетка - 4 n и т. д. Таким образом, результат эндомитоза полиплоидия. Пло́идность — число одинаковых наборов хромосом, находящихся в ядре клетки или в ядрах клеток многоклеточного организма. Диплоидные клетки имеют две одинаковые копии каждой хромосомы. Полиплоиди́ей -называют кратное увеличение количества хромосом в клетке эукариот. Различают автополиплоидию и аллополиплоидию. А́втополиплоиди́я — наследственное изменение, кратное увеличение числа наборов хромосом в клетках организма одного и того же биологического вида. На основе искусственной автополиплоидии синтезированы новые формы и сорта ржи, гречихи, сахарной свёклы и других растений. А́ллополиплоиди́я — кратное увеличение количества хромосом у гибридных организмов. Возникает при межвидовой и межродовой гибридизации. 22. Понятие о жизненном цикле клеток. Особенности жизненного цикла у различных видов клеток. Жизненный (клеточный) цикл - это весь период существования клетки (от деления до смерти или следующего деления). Клеточный цикл состоит из митотического периода (М) и интерфазы (межмитотического периода). Интерфаза в свою очередь состоит из пресинтетического (G 1), синтетического (S) и постсинтетического (G2) периодов. В пресинтетическом (постмитотическом, G1) периоде дочерняя клетка достигает размеров и структуры материнской, для чего в ней происходит биосинтез РНК и белков цитоплазмы и ядра. Кроме того, в ней синтезируются РНК и белки, Необходимые для синтеза ДНК в следующем периоде. В Синтетическом (S) периоде происходит удвоение (редупликация) ДНК и, соответственно, удваивается число хромосом (их количество становится тетраплоидным, 4n). В постсинтетическом (премитотическом, G2)периоде клетка готовится к митозу, в ней происходит синтез РНК и белков (тубулинов) веретена деления, накопление энергии, необходимой для митоза. Вышеописанный жизненный цикл характерен для популяции клеток, которые непрерывно делятся. Кроме того, в организме есть клетки, которые временно или постоянно находятся вне митотического цикла (в G0 периоде). Этот период характеризуется как состояние репродуктивного покоя. 23. Внутриклеточная регенерация. Регенерация внутриклеточная — это Р. поврежденных органоидов и мембранных структур клетки; характерна для клеток миокарда, нервной системы. Внутриклеточная форма регенерации является универсальной, так как она свойственна всем органам и тканям без исключения. Однако структурно-функциональная специализация органов и тканей в фило- и онтогенезе «отобрала» для одних преимущественно клеточную форму, для других — преимущественно или исключительно внутриклеточную, для третьих — в равной мере обе формы регенерации. К органам и тканям, в которых преобладает клеточная форма регенерации, относятся кости, эпителий кожи, слизистые оболочки, кроветворная и рыхлая соединительная ткань и т. д. Клеточная и внутриклеточная формы регенерации наблюдаются в железистых органах (печень, почка, поджелудочная железа, эндокринная система), легких, гладких мышцах, вегетативной нервной системе. К органам и тканям, где преобладает внутриклеточная форма регенерации, относятся миокард и скелетные мышцы, в центральной нервной системе эта форма регенерации становится единственной формой восстановления структуры. Преобладание той или иной формы регенерации в определенных органах и тканях определяется их функциональным назначением, структурно-функциональной специализацией. 24. Апоптоз и его биологическое значение. Апоптоз– это контролируемый физиологический процесс самоуничтожения клетки, характеризующийся поэтапным разрушением и фрагментацией ее содержимого с формированием мембранных пузырьков (апоптозных телец), впоследствии поглощаемых фагоцитами. Этот генетически заложенный механизм активируется под воздействием определенных внутренних или внешних факторов. Биологическое значение апоптоза заключается в следующем: 1. Осуществление нормального развития организма в период эмбриогенеза. 2. Предотвращение размножения мутировавших клеток. 3. Регуляция деятельности иммунной системы. 4. Предотвращение преждевременного старения организма Данный процесс играет ведущую роль в эмбриогенезе, поскольку многие органы и ткани претерпевают значительные трансформации во время эмбрионального развития. Многие врожденные дефекты возникают вследствие недостаточной активности апоптоза. Как запрограммированная самоликвидация дефектных клеток, данный процесс является мощной природной защитой против онкологических заболеваний. Так, к примеру, вирус папилломы человека блокирует клеточные рецепторы, ответственные за апоптоз и, таким образом, приводит к развитию рака шейки матки и некоторых других органов. Благодаря данному процессу происходит физиологическая регуляция клонов Т-лимфоцитов, ответственных за клеточный иммунитет организма. Клетки, неспособные распознавать белки собственного организма (а таких в общей сложности созревает около 97%), подвергаются апоптозу. Недостаточность апоптоза приводит к тяжелым аутоиммунным заболеваниям, в то время как его усиление возможно при иммунодефицитных состояниях. К примеру, тяжесть течения СПИДа коррелирует с усилением данного процесса у Т-лимфоцитов. 25. Современный этап в развитии гистологии и цитологии. Развитие методов гистологических и цитологических исследований. Метод культивирования тканей в организме по Ф.М. Лазаренко. Развитие гистологии. Успехи гистологии как науки о строении и происхождении тканей и их компонентов, прежде всего, связаны с развитием техники, оптики и методов микроскопирования. Микроскопические исследования позволили накопить данные по тонкому строению организма и на этом основании сделать теоретические обобщения. В истории учения о тканях и микроскопическом строении органов следует различать три периода: 1й — домикроскопический (продолжительностью около 2000 лет), 2-й — микроскопический (около 300 лет), 3й — современный, сочетающий достижения в области электронной микроскопии, иммуноцитохимии, цитофотометрии и др. (с середины XX столетия). Методы гистологических и цитологических исследований Световая микроскопия Электронная микроскопия Цитои гистохимия Иммуноцитохимия Авторадиография Методы молекулярной биологии Культура тканей и клеток Клеточная инженерия ЛАЗАРЕНКО МЕТОД- метод имплантации измельченных тканей животного организма, применяемый в эксперименте для изучения особенностей жизнедеятельности тканей. Опубликован Ф. М. Лазаренко в 1934 г. Ткани того или иного органа животного в асептических условиях измельчают на кусочки размером до 2/3 мм3, смешивают с мелкими кусочками нейтрального целлоидина в соотношении 1:2 и имплантируют в подкожную клетчатку животномуреципиенту. В области имплантата образуется очаг асептического воспаления, вокруг к-рого образуется соединительнотканная капсула. В имплантат врастают кровеносные сосуды и нервы реципиента, обеспечивающие трофику имплантируемых тканей. Культивируемые ткани проходят ряд последовательно сменяемых состояний: депрессия, пролиферация, органогенез, дифференцировка и обратное развитие новообразованных в имплантате структур. Л. м. может быть использован для изучения реактивных и пластических свойств тканей (преимущественно эпителиальных), влияний регуляторных систем организма на тканевые процессы и их дифференцировку, для экспериментального моделирования процессов органогенеза 2. Эмбриология 26. Понятие прогенеза и эмбриогенеза. Особенности структуры половых клеток человека. Прогенез - это период развития и созревания половых клеток — яйцеклеток и сперматозоидов. В результате прогенеза в зрелых половых клетках возникает гаплоидный набор хромосом, формируются структуры, обеспечивающие их способность к оплодотворению и развитию нового организма. Подробно процесс развития половых клеток рассмотрен в главах, посвященных мужской и женской половым системам. Эмбриогенез человека — часть его онтогенеза, включающая следующие основные стадии: I — оплодотворение, и образование зиготы; II — дробление и образование бластулы (бластоцисты); III — гаструляцию — образование зародышевых листков и комплекса осевых органов; IV — гистогенез и органогенез зародышевых и внезародышевых органов; V — системогенез. 27. Сперматогенез. Стадии сперматогенеза: размножение; роста; созревание-деление; формирование. Начальной фазой сперматогенеза является размножение сперматогоний путем митоза, большая часть клеток продолжает делится, а меньшая часть вступает в стадию роста. В этот период клетки растут, накапливают питательные вещества, и потом превращаются в сперматоциты 1-го порядка. Следующая фаза созревание-деление, характеризуется двумя редукционными делениями, без интерфазы. В результате 1-го деления 1 сперматоцит 1-го порядка дает начало 2м сперматоцитам 2-го порядка, а 2-ое деление-созревание приводит к появлению 4 сперматид. Фаза формирования происходит в присутствии тестостерона, происходит преобразование сперматид в сперматозоиды. Сперматозоиды — это мелкие, подвижные клетки, размером 30—60 мкм. В сперматозоиде различают головку и хвост. Головка сперматозоида имеет овоидную форму и включает в себя небольшое плотное ядро, окруженное тонким слоем цитоплазмы. Ядра сперматозоидов характеризуются высоким содержанием нуклеопротаминов и нуклеогистионов. Передняя половина ядра покрыта плоским мешочком, составляющим "чехлик" сперматозоида. В нем у переднего полюса располагается акросома. Чехлик и акросома являются производными комплекса Гольджи. Акросома содержит набор ферментов, среди которых важное место принадлежит гиалуронидазе и протеазам, способным растворять оболочки, покрывающие яйцеклетку. За головкой имеется кольцевидное сужение. Головка так же, как и хвостовой отдел, покрыта клеточной мембраной. Хвостовой отдел сперматозоида состоит из связующих, промежуточных, главной и терминальной частей. В связующей части или шейке располагаются центриоли — проксимальная и дистальная, от которой начинается осевая нить (аксонема). Промежуточная часть содержит 2 центральных и 9 пар периферических микротрубочек, окруженных расположенными по спирали митохондриями. Именно митохондрии обеспечивают энергией двигательную активность сперматозоидов, нарушение которой нередко связано с поражением процесса энергообразования в митохондриях. Главная часть по строению напоминает ресничку. Она окружена тонким фибриллярным влагалищем. Терминальная, или конечная часть содержит единичные сократительные филаменты. 28. Овогенез. Овогенез — это процесс образования и развития женских половых клеток. Он включает в себя 3 фазы: размножения; роста; созревания. Фаза размножения начинается в эмбриональном периоде и продолжается в течение 1-го года жизни девочки. К моменту рождения у девочки имеется около 2-х млн клеток. К периоду полового созревания остается около 40 тыс. половых клеток и в последующем 1 раз в 28—32 дня происходит созревание и выход одной яйцеклетки в маточную трубу — овуляция. Овуляция прекращается при наступлении беременности или менопаузы. Сущностью фазы размножения является митотическое деление овогоний. Фаза роста, в конце 1-го года жизни девочки размножение овогоний останавливается и клетки яичника вступают в фазу малого роста, превращаясь в овоциты 1-го порядка. Наступает 1 блок роста, который снимается с наступлением полового созревания, то есть появлением женских половых гормонов. Далее овоциты 1-го порядка вступают в фазу большого роста. Фаза созревания, как и во время сперматогенеза, включает в себя два деления, причем второе следует за первым без интеркинеза, что приводит к уменьшению (редукции) числа хромосом вдвое, и набор из становится гаплоидным. При первом делении созревания овоцит 1-го порядка делится, в результате чего образуются овоцит 2-го порядка и небольшое редукционное тельце. Овоцит 2-го порядка получает почти всю массу накопленного желтка и поэтому остается столь же крупным по объему, как и овоцит 1-го порядка. Редукционное же тельце представляет собой мелкую клетку с небольшим количеством цитоплазмы. При втором делении созревания в результате деления овоцита 2-го порядка образуются одна яйцеклетка и второе редукционное тельце. Первое редукционное тельце иногда тоже делится на две одинаковые мелкие клетки. В результате этих преобразований овоцита 1-го порядка образуются одна яйцеклетка и три редукционных тельца. 29. Оплодотворение у человека. Биологическое значение оплодотворения, особенности и хронология процесса. Зигота, её геном, активация внутриклеточных процессов. Оплодотворение — процесс слияния мужской и женской гамет, приводящее к образованию зиготы. При оплодотворении взаимодействуют мужская и женская гаплоидные гаметы, при этом сливаются их ядра (пронуклеусы), объединяются хромосомы, и возникает первая диплоидная клетка нового организма — зигота. Начало оплодотворения — момент слияния мембран сперматозоида и яйцеклетки, окончание оплодотворения — момент объединения материала мужского и женского пронуклеусов. Оплодотворение происходит в дистальном отделе маточной трубы и проходит 3 стадии: I стадия — дистантное взаимодействие, включает в себя 3 механизма: хемотаксис — направленное движение сперматозидов навстречу к яйцеклетке (гинигамоны 1,2); реотаксис — движение сперматозоидов в половых путях против тока жидкости; капацитация — усиление двигательной активности сперматозоидов, под воздействием факторов женского организма (рН, слизь и другие). II стадия — контактное взаимодействие, за 1,5—2 ч сперматозоиды приближаются к яйцеклетке, окружают ее и приводят к вращательным движениям, со скоростью 4 оборота в минуту. Одновременно из акросомы сперматозоидов выделяются сперматозилины, которые разрыхляют оболочки яйцеклетки. В том месте где оболочка яйцеклетки истончается максимально происходит оплодотворение, оволемма выпячивается и головка сперматозоида проникает в цитоплазму яйцеклетки, занося с собой центриоли, но оставляя снаружи хвостик. III стадия — проникновение, самый активный сперматозоид приникает головкой в яйцеклетку, сразу после этого в цитоплазме яйцеклетки образуется оболочка оплодотворения, которая препятствует полиспермии. Затем происходит слияние мужского и женского пронуклеусов, этот процесс носит название синкарион. Этот процесс (сингамия) и есть собственно оплодотворение, появляется диплоидная зигота (новый организм, пока одноклеточный 30. Первая неделя развития зародыша человека. Специфика дробления у человека и хронология процесса. Строение зародыша перед имплантацией. Дробление — это последовательно протекающий митоз, без роста образовавшихся клеток, до размеров исходной. При дроблении происходит относительно быстрое увеличение количества клеток (бластомеры). Дробление идет до тех пор, пока не восстановится соотношение объема ядра к объему цитоплазмы, характерное для данного вида. Количество бластомеров увеличивается от 2 до примерно 12—16 к третьим суткам после оплодотворения, когда концептус достигает стадии морулы и выходит в полость матки из маточных труб. Различают дробление: полное, неполное; равномерное, неравномерное; синхронное, асинхронное. У человека дробление полное, асинхронное, неравномерное. В результате первого деления образуются 2 бластомера, темный и светлый, светлые делятся быстро и обволакивают зиготу снаружи — трофобласт, а темные находятся внутри и делятся медленно — эмбриобласт. Дробление зиготы у человека прекращается на стадии 107 бластомеров.. 31. Имплантация. Хронология процесса имплантации. Дифференцировка трофобласта на цитотрофобласт и синцитиотрофобласт. Имплантация состоит из 2-х этапов: адгезияприлипание; инвазияпогружение. На 4-е сутки после оплодотворения, в полость матки выпадает морула. Морула — группа клеток, возникших в ходе нескольких делений дробления и заключенных внутри прозрачной оболочки. Около 2-х суток морула находится в полости матки в неприкрепленном состоянии, при этом клетки трофобласта поглощают из окружающей среды питательные вещества и воду, жидкость накапливается в моруле и она превращается в бластоцисту. Бластоциста возникает с появлением бластоцеля (заполненной жидкостью полости), объем бластоцеля увеличивается и зародыш приобретает форму пузырька. Прозрачная оболочка истончается и исчезает. Одновременно с процессом имплантации, в зародыше начинается гаструляция. Сущностью процесса является законченное перемещение бластомеров с образованием 3-х зародышевых листков. У млекопитающих в гаструляции различают следующие процессы: инвагинацию — вдавление; эпиболию — обрастание; эмиграцию — выселение, перемещение; деляминацию — расщепление. Двухслойный зародыш начинает дальше дифференцироваться на светлые и темные бластомеры. Из крупных, более светлых клеток, составляющих наружный слой, развивается трофобласт и другие внезародышевые образования (это среда обитания и обеспечения). Более темные и мелкие клетки дают начало развития эмбриобласту (формирование трехзародышевых листков). В дальнейшем трофобласт дифференцируется на цитотрофобласт и синцитиотрофобласт. Синцитиотрофобласт состоит из больших многоядерных клеток, развивающихся из цитотрофобласта и обладающих инвазивными свойствами. Синцитиотрофобласт внедряется в строму эндометрия, разрушает кровеносные сосуды и формирует сосудистые лакуны. Затем синцитиотрофобласт достигает спиральных артерий эндометрия. 32. Гаструляция у человека. Гаструляция – сложный процесс химических и морфогенетических изменений, сопровождающийся размножением, ростом, направленным перемещением и дифференцировкой клеток, в результате образуются зародышевые листки: наружный – эктодерма, внутренний – энтодерма, средний – мезодерма. Это источники зачатков тканей и органов, комплекса осевых органов. Способы гаструляции: Инвагинация (впячивание) – дно подтягивается к крыше Эпиболия (обрастание) – характерна для крупных клеток, переполненных желтком. Иммиграция (выселение) Деляминация (расщепление) У человека протекает в две фазы: Деляминация – зародышевый щиток (эмбриобласт) расщепляется на эпибласт (все зачатки кроме энтодермы) и гипобласт (энтодерма). ИММИГРАЦИЯ (на 14-17 сутки) – движение клеточных масс от головного и хвостового концов, затем они идут параллельно, образуя первичную полоску. Материал, который дремал в головном конце приходит в движение и идет навстречу первому потоку. Клеточный материал перестраивается и образуется первичный узелок с ямкой в центре. Через первичный (Гензеновский) узелок перемещается прехордальный зачаток и уходит в энтодерму (в головной конец зародыша); хордальный зачаток через первичный узелок ложится между экто- и энтодермой, образуя хорду. Мезодерма двумя крыльями подворачивается через края первичной полоски и уходит внутрь, располагаясь по бокам от хорды. Эктодерма всегда перемещается пластом, мезодерма – совершает амебообразные движения, энтодерма относительно неподвижна. Факторы вызывающие гаструляцию: Неравномерный рост в различных областях зародыша Механический (бластомеры легко изменяют форму) Разная интенсивность поглощения воды Различная пластичность клеточных мембран Способность клеток к фагоцитозу Межклеточные взаимодействия 33. Вторая неделя развития человека. Строение двухнедельного зародыша. К концу второй недели (14 сутки) образуется второй пузырек – желточный. К этому времени внезародышевая мезодерма расслаивается на париетальный (прилегает трофобласту) и висцеральный листки. Амниотическая ножка трофобласта превращается в хорион – это трофобласт + внезародышевая мезодерма. Из мезодермы образуется мезенхима, которая врастает в первичные ворсинки трофобласта. Так образуются вторичные ворсинки, содержащие кровеносные сосуды. Сам материал будущего плода располагается в области дна амниотического и крыши желточного пузырей. Т.о. к концу второй недели зародыш полностью имплантирован, трофобласт преобразовался в хорион, есть амниотический, желточный пузырьки. Материал самого зародыша располагается в области дна амниотического пузырька и в области крыши желточного пузырька. Дно амниотического пузырька представляет собой эпибласт, а крыша желточного – гипобласт. Они делятся путем деляминации, и с этого времени начинается вторая фаза гаструляции. 34. Представление о критических периодах развития. (Н. Грэгг, П. Г. Светлов). 1 . Прогенез или гаметогенез , который характеризуется специфическими изменениями числа хромосом в мейозе . 2 . Оплодотворение - слияние гамет и восстановление диплоидной числа хромосом . 3 . Имплантация - врастание эмбриона в эндометрий (7-8 день). 4 . Гаструляция , нейруляция и образование комплекса осевых зачатков ( С - 8 неделя ) . 5 . Усиленный рост головного мозга (15-20 недели) . 6 . Органо - и системогенез (формирование жизненно важные системы 20 по 24 неделю). 7 . Рождение . 8 . Период новорожденности и первый год жизни. 9 . Половое дозривння (11-16 лет). 10 . Менопауза . 35. Дифференцировка зародышевых листков и образование зачатков и органов у зародыша человека. Факторы, вызывающие дифференцировку. Дифференцировка первичной эктодермы: Зародышевая эктодерма Нервная трубка (нейроциты и нейроглия сетчатки глаза и органа обоняния, нейроциты и нейроглия головного и спинного мозга) Нервный гребень и ганглиозные пластинки (нейроциты и нейроглия спинальных и вегетативных ганглиев, хромаффинная ткань и мозговое в-во надпочечников) Плакоды (эпителиальные элементы внутреннего уха) Кожная эктодерма (эпидермис и его производные, эпителий роговицы глаза, эпителий органов ротовой полости и его производные, эпителий анального отдела прямой кишки, эмаль и кутикула зуба, эпителиальная выстилка влагалища) Прехордальная пластинка (эпителий ротовой полости и пищевода, эпителий трахеи, бронхов и легких) Внезародышевая эктодерма (эпителий амниона и пупочного канатика) Дифференцировка первичной энтодермы Зародышевая энтодерма Энтодерма кишечной трубки (эпителий кишечника, желудка, печени,поджелудочной) Внезародышевая энтодерма Желточная энтодерма (эпителий аллантоиса и желточного места) Дифференцировка мезодермы Зародышевая мезодерма Сомиты Миотом (скелетная мышечная ткань) Склеротом (хрящевая и костная ткань) Дерматом (соединительнотканная основа кожи) Сегментная ножка нефрогонотома (эпителий гонад и семявыносящих путей и почек) Парамезонефральный проток (эпителиальная выстилка влагалища, матки, яйцеводов) Спланхнотом (поперечнополосатая мышечная ткань сердца, корковое в-во надпочечников, мезотелий) Мезенхима (микроглия, гладкая мышечная ткань, сосуды, соединительные ткани, к-ки крови и кроветворных органов) Внезародышевая мезодерма Мезенхима (экзоцелломический эпителий, соединительная ткань желточного мешка амниона и хориона) Спланхнотом Париетальный и висцеральный листки 36. Амнион, его образование, развитие, строение, функции. Амнион — временный орган, обеспечивающий водную среду для развития зародыша. Он возник в эволюции в связи с выходом позвоночных животных из воды на сушу. В эмбриогенезе человека он появляется на второй стадии гаструляции сначала как небольшой пузырек, дном которого является первичная эктодерма (эпибласт) зародыша. Стенка пузырька образует внезародышевую эктодерму, которая соединяется с внезародышевой мезодермой, разрастается и окружает зародыш тонкой полупрозрачной амниотической оболочкой (источник развития его эпителия) Амниотическая оболочка образует стенку резервуара, заполненного амниотической жидкостью, в которой находится плод. Основная функция амниотической оболочки выработка околоплодных вод, обеспечивающих среду для развивающегося организма и предохраняющих его от механического повреждения. Эпителий амниона, обращенный в его полость, не только выделяет околоплодные воды, но и принимает участие в обратном всасывании их. В амниотической жидкости поддерживаются до конца беременности необходимый состав и концентрация солей. Амнион выполняет также защитную функцию, предупреждая попадание в плод вредоносных агентов. 37. Желточный мешок, его образование, развитие, строение, функции. Из краев гипобласта выселяются клетки внезародышевой энтодермы и подрастают у ранее образовавшейся мезенхимальной закладки - желточного мешка. Желточный мешок — вынесенная за пределы зародыша часть первичной кишки. Стенка желточного мешка состоит из двух слоев. Внутренний слой образован внезародышевой энтодермой, а наружный — внезародышевой мезодермой. Складки амниона сдавливают желточный мешок, образуя узкую перемычку, соединяющую его с полостью первичной кишки — желточный стебелек. Эта структура удлиняется и вступает в контакт ножкой тела, содержащей аллантоис. Желточный мешок обычно полностью зарастает к концу 3-го месяца развития плода. Задняя стенка желточного мешка к 14—16-му дню развития формирует небольшой вырост — аллантоис, образованный внезародышевыми энтодермой и мезодермой. Дистальная часть аллантоиса по мере роста быстро расширяется и превращается в мешок, соединенный с кишкой при помощи ножки. У человека аллантоис рудиментарно участвует в формировании сосудистой сети плаценты. Его проксимальный отдел имеет отношение к образованию мочевого пузыря, что следует учитывать при аномалиях развития этого органа. Развитие, строение и функции желточного мешка. Из гипобласта выселяются клетки, составляющие внезародышевую (или желточную) энтодерму, и, обрастая изнутри мезенхимальную закладку желточного мешка, образуют вместе с ней стенку желточного мешка. Стенка желточного мешка состоит из: 1) внезародышевой (желточной) энтодермы;2) внезародышевой мезенхимы. Функции желточного мешка: 1) кроветворение (образование стволовых клеток крови);2) образование половых стволовых клеток (гонобластов);3) трофическая (у птиц и рыб). 38. Аллантоис, его образование, развитие, строение, функции. Аллантоис развивается на 16-17-е сутки в виде небольшого выроста задней стенки желточного мешка, поэтому имеет те же оболочки, что и желточный мешок: внезародышевая энтодерма изнутри и внезародышевая мезодерма снаружи. Аллантоис врастает в амниотическую ножку, в его стенке развиваются пупочные кровеносные сосуды, которые он подводит к хориону. Таким образом, аллантоис выполняет ту же функцию, что и регрессирующий желточный мешок – они играют роль проводников и направляют рост сосудов плода к плаценте. На втором месяце эмбрионального развития аллантоис редуцируется и вместе с остатками желточного мешка образует клеточный тяж в составе пупочного канатика. Кроме того, аллантоис участвует в развитии мочевого пузыря. Развитие, строение и функции аллантоиса. Часть зародышевой энтодермы гипобласта в виде пальцевидного выпячивания врастает в мезенхиму амниотической ножки и формирует аллантоис. Стенка аллантоиса состоит из: 1) зародышевой энтодермы; 2) внезародышевой мезенхимы. Функциональная роль аллантоиса: 1) у птиц полость аллантоиса достигает значительного развития и в ней накапливается мочевина, поэтому его называют мочевым мешком; 2) у человека нет необходимости накопления мочевины, поэтому полость аллантоиса очень незначительная и к концу 2-го месяца полностью зарастает. 39. Хорион, его образование, развитие, строение, функции. Первым из провизорных органов образуется хорион.Развитие хориона начинается с выселения клеток внезародышевой мезенхимы из зародышевого щитка.Часть этих клеток изнутри подрастает к цитотрофобласту. С этого момента трофобласт становится трёхслойным (симпластотрофобласт, цитотрофобласт, внезародышевая мезенхима) и носит название хориона. На второй неделе эмбриогенеза на поверхности хориона формируются первичные ворсины – выросты, состоящие из цитотрофобласта, расположенного в центре, и симпластотрофобласта, размещённого на поверхности ворсины. Когда в ворсины изнутри врастает внезародышевая мезенхима, они становятся вторичными. На 3-ейнеделе эмбриогенеза во вторичных ворсинах, расположенных в области прикрепления к хориону амниотической ножки, формируются первые клетки крови и кровеносные сосуды. С этого момента ворсины называются третичными. Третичные ворсины разрастаются, разветвляются, и образуется ворсинчатый хорион. На остальной поверхности хориона ворсины редуцируются. Эта часть хориона называется гладким хорионом. Из ворсинчатого хориона развивается плодная часть плаценты, гладкий хорион входит в состав плодного яйца. Хорион выполняет все функции, присущие плаценте – барьерная, защитная, трофическая, секреторная, эндокринная. 40.Третья неделя эмбриогенеза человека. Строение трёхнедельного зародыша. В этот период продолжается вторая фаза гаструляции, образуются зародышевые листки, хорда, прехордальная пластинка, нервная трубка, нервный гребень. Начинается сегментация дорсальной мезодермы (сомиты, сегментные ножки), образуются париетальный и висцеральный листки спланхнотома и эмбриональный целом, который в дальнейшем разделяется на три полости тела: 1перикардиальную;2плевральную;3перитонеальную; Закладываются сердце, кровеносные сосуды, предпочка (пронефрос). Происходит формирование внезародышевых органов – аллантоиса, вторичных и третичных ворсин хориона. Образуется туловищная складка. Первичная кишка зародыша отделяется от вторичного желточного мешка. Обособление тела зародыша С 20–х – 21–х суток начинает обособляться тело зародыша Формирование комплекса осевых зачатков В течение конца 3–й и всей 4–й недель (18–28 сутки) из трёх зародышевых листков формируется комплекс осевых зачатков . В свою очередь, затем из большинства этих зачатков развиваются ткани, органы и системы. Дифференцировка мезодермы и образование мезенхимы. Сразу после своего образования мезодерма подразделяется на два главных отдела: сомиты – спинной отдел и спланхнотом – брюшной отдел. Между сомитами и спланхнотомом имеется еще один отдел – сегментная ножка, с помощью которой они соединяются. Сомиты разделяются на три части: дерматом, склеротом, миотом. Спланхнотом делится на висцеральный и париетальный листки, между которыми находится вторичная полость тела – целом. Висцеральный и париетальный листки дают начало висцеральным и париетальным серозным оболочкам. Образование хорды происходит из клеток первичного узелка. Клетки первичного узелка, образующегося в эктодерме, прорастают в пространство между экто– и энтодермой, там разрастаются вперед и назад и образуют хорду. Хорда образуется практически одновременно с самой мезодермой – в конце 3–й недели развития. Вначале часть клеток эпибласта, мигрирующих через первичную ямку, формирует отросток – нотохорд. Последний растёт по оси зародыша (между крыльями мезодермы) вперёд – пока не достигает прехордальной пластинки. В итоге, образуется непарная осевая структура (хорда), вокруг которой впоследствии формируется позвоночный столб. Во взрослом организме хорда сохраняется в виде пульпозных ядер межпозвонковых дисков. Образование нервной трубки. Уже после образования мезодермы и хорды, под индуктивным влиянием нотохорда (растущего вперёд) в срединной части эктодермы образуется утолщение, тоже растущее вперёд от первичного узелка, – нервная пластинка. На 18–й день начинается прогибание этой пластинки – появляются нервный желобок и нервные валики. Затем ( на протяжении 4–й недели) желобок постепенно замыкается: вначале – в шейном отделе, потом – в каудальном и, наконец, в головном – образуется непарная нервная трубка. Образование кишечной трубки происходит в процессе формирования туловищной складки. Туловищная складка формируется путем подгибания всех имеющихся зародышевых листков под тело зародыша. В результате этого кишечная (зародышевая) энтодерма отделяется от желточного мешка (желточная энтодерма). Кишечная трубка остается связанной с желточным мешком только в одном небольшом участке – желточном протоке (стебельке). Часть желточного протока может остаться после рождения в виде дивертикула подвздошной кишки. 41. Плацента. Типы плацент. Плацента — место связи аллантохориона плода со слизистой оболочкой матки. Она образуется за счет аллантохорионной оболочки и слизистой оболочки матки. В соответствии с этим в ней различают детскую и материнскую части. Различают: зародышевую часть: ворсинки хориона материнскую часть: слизистая матки, где имплантировался зародыш. Морфологическая классификаци: диффузная: образование ворсин по всей поверхности хориона (свиньи); островная (котиледонная): ворсинки хориона собраны в группы (котиледоны), между которыми поверхность хориона гладкая (жвачные); поясная (зонарная): ворсинки хориона сосредоточены внутри небольшого участка, опоясывающего плодный пузырь (хищные). дискоидальная: вид диска, ворсинки хориона распределены по всей ее поверхности равномерно (у человекообразных обезьян и человека, у мартышек и макак – бидискоидальная). В норме у всех млекопитающих кровь зародыша и кровь матери нигде не смешиваются (отделены стенками сосудов зародыша и тканями хориона). Гистологическая классификация: по глубине погружения ворсинок хориона зародыша и степени их проникновения в слизистую оболочку матки : эпителиохориальная: наиболее примитивная (сумчатые, парнокопытные, китообразные). Ворсинки хориона не прободают даже эпителиальной выстилки матки, а лишь погружаются в складки ее слизистой оболочки. Нет контакта между ворсинками хориона и материнской частью десмохориальная (соединительно-тканная): ворсинки хориона в месте контакта разрушают слизистую оболочку матки и внедряются в ее соединительнотканный слой, контакт по типу десмосом (жвачные). эндотелиохориальная: устанавливается контакт между сосудами плода и матери. Ворсинки хориона отделены от сосудов матери лишь эндотелиальной стенкой (хищные). гемохориальная: тесная связь сосудов плода и матери (приматы, грызуны). Ворсинки хориона погружаются в кровяные лакуны, заполненные кровью матери. 42. Плацентация у человека. Строение сформированной плаценты. Развитие и строение плаценты человека. Плацента че­ловека начинает развиваться на 3-й и заканчивается на 6-8-й неделе (плацентация). Окончательно плацента форми­руется на 12-й неделе. Плацента состоит из 2 частей: плод­ной (pars fetalis) и маточной (pars materna). Плодная часть плаценты развивается из ветвистого хориона. Ворсины ветвистого хориона погружаются в базальную отпадающую оболочку эндометрия матки, в результате чего в этой оболочке образуются углу­бления — лакуны. В лакунах циркулирует материнская кровь. Базальная отпадающая оболочка с лакунами — это даточная часть плаценты. Строение плодной части плаценты на 12-й неделе. Плодная часть плаценты включает: 1) хориальную пластин­ку, состоящую из внезародышевой мезодермы (соедини­тельной ткани); 2) амниотическую оболочку, которая сра­стается с внутренней поверхностью хориальной пластинки; 3) цитотро- и синцитиотрофобласт, покрывающие наруж­ную поверхность хориальной пластинки, обращенной к эн­дометрию; 4) третичные ворсины, погруженные в лакуны. Третичные ворсины отходят от наружной, поверхности хо­риальной пластинки. От каждой такой ворсины отходят ветви. От основного ствола ворсины отходят вторичные ве­тви (ворсины), от вторичных — третичные. Основу ворсин образует соединительная ткань (соедини­тельнотканная строма). Эта строма покрыта цитотрофобластом, лежащим на базальной мембране, и синцитиотрофобластом, расположенным снаружи ворсины. На поверхности синцитиотрофобласта имеются микроворсинки, которые со­вокупности образуют всасывающую каемку. Каждая третич­ная ворсина представляет собой котиледон. Таких котиледо­нов в плодной части плаценты около 200. В строме ворсины проходят кровеносные сосуды, в которых циркулирует кровь плода. Среди ворсин имеются «якорные» ворсины. Эти ворсины характеризуются тем, что они при помощи периферического цитотрофобласта прикрепляются к маточной части плаценты. В трофобласте ворсин содержится около 60 различных ферментов: СДГ, цитохромоксидаза, ЩФ, кислая фосфатаза, АТФаза, глюкозо-6-дегидрогеназа и др. При помощи этих ферментов питательные вещества, всосавшиеся в трофобласт из крови матери, расщепляются до простейших соеди­нений. Белки распадаются до аминокислот. Из этих амино­кислот тут же, в трофобласте, синтезируются специфичные для плода белки. Готовые белки затем поступают в капилля­ры ворсин, в которых циркулирует кровь плода. 43. Система “мать-плод”. Гематоплацентарный барьер. Система мать-плод возникает в процессе беременности и включает две подсистемы - организм матери и организм плода, а также плаценту, являющуюся связующим звеном между ними. Взаимодействие между организмом матери и организмом плода обеспечивается прежде всего нейрогуморальными механизмами. При этом в обеих подсистемах различают следующие механизмы: рецепторные, воспринимающие информацию, регуляторные, осуществляющие ее переработку, и исполнительные. Рецепторные механизмы организма матери расположены в матке в виде чувствительных нервных окончаний, которые первыми воспринимают информацию о состоянии развивающегося плода. В эндометрии находятся хемо-, механо- и терморецепторы, а в кровеносных сосудах - барорецепторы. Рецепторные нервные окончания свободного типа особенно многочисленны в стенках маточной вены и в децидуальной оболочке в области прикрепления плаценты. Раздражение рецепторов матки вызывает изменения интенсивности дыхания, кровяного давления в организме матери, что обеспечивает нормальные условия для развивающегося плода. Гематоплацентарный барьер включает все структуры третичной ворсины хориона плода: эндотелий капилляра соматического типа, базальная мембрана капилляра, соединительнотканная строма ворсинки, базальная мембрана трофобласта, внутренний слой трофобласта - клетки цитотрофобласта , наружный слой трофобласта симпластотрофобласт (синцитиотрофобласт), который непосредственно контактирует с материнской кровью, находящейся в лакунах эндометрия матки. Ядра симпластотрофобласта овальные, с гетерохроматином по периферии. В базальной части симпластотрофобласта находятся хорошо развитые органеллы: аЭПС и грЭПС, комплекс Гольджи митохондрии , в апикальной части – множество секреторных гранул , пиноцитозных пузырьков , на поверхности имеются микроворсинки , по строению и функциям подобные микроворсинкам столбчатых энтероцитов. Щёточная каёмка принимает участие в транспорте веществ. 44. Взаимоотношения зародыша 9,5 недель, внезародышевых органов и оболочек матки. Важная роль в развитии зародыша позвоночных принадлежит внезародышевым оболочкам, или провизорным органам. Они являются временными органами и у взрослого организма отсутствуют. Провизорные органы обеспечивают важнейшие функции развивающегося зародыша, однако в состав его тела не входят, являясь тем самым внезародышевыми органами. К ним относятся желточный мешок, амнион, хорион, аллантоис и плацента. Динамика взаимоотношений зародыша, внезародышевых органов и оболочек матки:а - зародыш человека 9,5 нед развития (микрофотография): 1 - амнион; 2 - хорион; 3 - формирующаяся плацента; 4 - пуповина Развитие внезародышевых органов у зародыша человека (схема): 1 - амниотический пузырек;1а - полость амниона;2 - тело эмбриона;3 - желточный мешок;4 - внеэмбриональный целом;5-первичные ворсины хориона;6 - вторичные ворсины хориона;7 - стебелек аллантоиса;8 - третичные ворсины хориона;9 - аллан-тоис;10 - пупочный канатик;11 - гладкий хорион;12 – котиледоны Хорион (или ворсинчатая оболочка) выполняет защитную и трофическую функции. Часть хориона (ворсинчатый хорион) внедряется в слизистую оболочку матки и входит в состав плаценты, которую иногда рассматривают как самостоятельный орган. Амнион (или водная оболочка) образуется только у наземных животных. Клетки амниона продуцируют амниотическую жидкость (околоплодные воды), в которой и развивается эмбрион, а затем – плод. После рождения ребенка хориальная и амниотическая оболочки отторгаются. Желточный мешок развивается в наибольшей степени у зародышей, образующихся из полилецитальных клеток, и потому содержит много желтка, откуда и происходит его название. Аллантоис – слепое выпячивание каудального конца кишечной трубки, окруженное внезародышевой мезенхимой. У животных, развивающихся в яйце, аллантоис достигает большого развития и выполняет функцию резервуара для продуктов обмена зародыша (главным образом мочевины). Именно поэтому аллантоис нередко называю мочевым мешком. У млекопитающих необходимость в накоплении продуктов обмена отсутствует, так как они поступают через маточно-плацентарный кровоток в организм матери и выводятся ее экскреторными органами. Поэтому у таких животных и человека аллантоис развит слабо и выполняет другие функции: в его стенке развиваются пупочные сосуды, которые разветвляются в плаценте и благодаря которым формируется плацентарный круг кровообращения. 20. Формирование комплекса осевых органов у человека. У млекопитающих имеется 4 осевых органа (хорда, нервная трубка, кишечная трубка, мезодерма). Образование осевых органов. Мезодерма образуется из клеток первичной полоски. Клетки первичной полоски, образовавшейся в эктодерме, прорастают в пространство между экто- и энтодермой и там разрастаются, образуя мезодерму. Образование хорды происходит из клеток первичного узелка. Клетки первичного узелка, образующегося в эктодерме, прорастают в пространство между экто- и энтодермой, там разрастаются вперед и назад и образуют хорду. Образование нервной трубки. В срединной части эктодермы образуется углубление — нервная пластинка. Нервная пластинка погружается под эктодерму, а края эктодермы смыкаются, и образуется нервная трубка. Образование кишечной трубки происходит в процессе формирования туловищной складки. Туловищная складка формируется путем подгибания всех имеющихся зародышевых листков под тело зародыша. В результате этого кишечная (зародышевая) энтодерма отделяется от желточного мешка (желточная энтодерма). Кишечная трубка остается связанной с желточным мешком только в одном небольшом участке — желточном стебельке (протоке). Из наружного зародышевого листка вычленяется материал нервной пластинки, которая сначала прогибается, образуя нервный желобок, а затем смыкается краями, превращаясь в нервную трубку - зачаток нервной системы. Эктодерма срастается над материалом нервной системы и в дальнейшем образует кожный эпителий. Хордальная пластинка сворачивается в круглый в сечении тяж - зачаток хорды. 21. Источники и ход эмбрионального развития органов нервной системы. Представляет собой систему тканей и органов, построенных из нервной ткани. В ней выделяют: Центральный отдел: головной и спинной мозг. Периферический отдел: автономные и чувствительные ганглии, периферические нервы, нервные окончания. Существует также подразделение на: Соматический (анимальный, цереброспинальный) отдел; Вегетативный (автономный) отдел: симпатическая и парасимпатическая части. Развитие. Нервная ткань развивается из дорсальной эктодермы. Стадия развития нервной трубки. У 18-дневного эмбриона человека эктодерма формирует нервную пластинку, латеральные края которой образуют нервные валики, а между валиками формируется нервный желобок. Передний конец нервной пластинки образует головной мозг. На 19ый день начинается процесс смыкания валиков над желобком, в результате чего образуется длинная трубка – нервная трубка. Она располагается под поверхностью эктодермы отдельно от нее. Клетки нервных валиков перераспределяются в один слой, в результате чего образуется ганглиозная пластинка. Из нее формируются все нервные узлы соматической периферической и вегетативной нервной системы. К 24-му дню развития трубка замыкается в головной части, а сутками позже – в каудальной. Клетки нервной трубки носят название медуллобластов. Клетки ганглиозной пластинки называются ганглиобластами. Медуллобласты затем дают начало нейробластам и спонгиобластам. Нейробласты отличаются от нейронов значительно меньшим размером, отсутствием дендритов, синаптических связей и вещества Ниссля в цитоплазме. Полость нервной трубки сохраняется у взрослых в виде системы желудочков головного мозга и центрального канала спинного мозга. Часть клеток нервной пластинки образует нервный гребень (ганглиозная пластинка). Стадия мозговых пузырей. В головном конце нервной трубки после ее замыкания очень быстро образуется три расширения – первичные мозговые пузыри. Полости первичных мозговых пузырей сохраняются в мозгу ребенка и взрослого в видоизмененной форме, образуя желудочки мозга и сильвиев водопровод. Существует две стадии мозговых пузырей: стадия трех пузырей и стадия пяти пузырей. Стадия формирования отделов мозга. Сначала образуются передний, средний и ромбовидный мозг. Затем из ромбовидного мозга образуются задний и продолговатый мозг, а из переднего образуются конечный мозг и промежуточный. Конечный мозг включает в себя два полушария и часть базальных ядер. В дальнейшем в нервной трубке дифференцируется 4 концентрических зоны: вентрикулярная (эпендимная), субвентрикулярная, промежуточная (плащевая) и краевая (маргинальная). 22. Источники и ход эмбрионального развития кровеносных сосудов и сердца. Сердечно-сосудистая система — совокупность органов (сердце, кровеносные и лимфатические сосуды), обеспечивающая распространение по организму крови и лимфы, содержащих питательные и биологически активные вещества, газы, продукты метаболизма. Кровеносные сосуды представляют собой систему замкнутых трубок различного диаметра, осуществляющих транспортную функцию, регуляцию кровоснабжения органов и обмен веществ между кровью и окружающими тканями. В эмбриональном развитии сердечно-сосудистой системы участвует мезенхима, висцеральные и париетальные листки спланхнотома. 1. Мезенхима. На 2-3 недели эмбриогенеза первые кровеносные сосуды появляются в мезенхиме желточного мешка и ворсинках хориона. Из мезенхимы на 17ые сутки справа и слева образуются эндокардиальные сердечные трубки , которые впячиваются в висцеральные листки спланхнотома. 2. Висцеральные листки спланхнотома. утолщённые участки спланхнотома – миоэпикардиальные пластинки, дадут начало миокарду и эпикарду. Из слившихся мезенхимных трубок формируется эндокард. Клетки миоэпикардиальных пластинок дифференцируются в 2х направлениях: из наружной части образуюется мезотелий, выстилающий эпикард. Клетки внутренней части дифференцируются в 3х направлениях. Из них образуются: сократительные кардиомиоциты; проводящие кардиомиоциты; эндокринные кардиомиоциты. 3. Париетальные листки спланхнотома . Из париетального листка спланхнотома развивается перикард. Перикард также выстлан мезотелием. В процессе развития сердца имеет место три этапа: 1)дифференцировка; 2)стадия стабилизации; 3)стадия инволюции. Дифференцировка начинается в эмбриогенезе и продолжается сразу после рождения. Стадия стабилизации начинается в двадцатилетнем возрасте и заканчивается в сорок лет. После сорока лет начинается стадия инволюции, сопровождаемая уменьшением толщины кардиомиоцитов засчёт уменьшения толщины миофибрилл. Увеличивается толщина прослоек соединительной ткани. Снижается частота и сила сокращений сердечной мышцы. Впоследствии это приводит к ишемической болезни сердца и инфаркту миокарда. 23. Источники и ход эмбрионального развития органов пищеварительной системы. Отделы и органы пищеварительной системы в эмбриогенезе развиваются из всех 3 зародышевых листков: эктодермы (эпителий ротовой и анальной ямок или бухт), энтодермы (эпителий первичной кишки или кишечной трубки) и мезодермы (собственная пластинка слизистой, подслизистая основа, мышечная, соединительнотканная и серозная оболочки). В конце 3-ей недели внутриутробного развития из энтодермы образуется трубка – первичная кишка, замкнутая на переднем и заднем концах. В конце 4-й недели внутриутробного развития на головном и каудальном концах эмбриона появляются углубления, которые соответственно называются ротовой и заднепроходной (анальной) бухтами (ямками). Образовавшиеся углубления отделяются от первичной кишки глоточной и анальной мембранами, состоящими из двух слоев: наружного – эктодермального и внутреннего – энтодермального. В первичной кишке выделяют глоточную и туловищную кишку. Границей между ними является выпячивание – закладка органов дыхания. В туловищной кишке в свою очередь выделяют три части: переднюю, среднюю и заднюю. Из различных отделов первичной кишки формируются следующие образования: Из глоточной кишки – задняя часть полости рта, язык, слюнные железы, небные миндалины, железы – производные эпителия глоточных карманов (щитовидная, околощитовидные, вилочковая). Из передней кишки –пищевод и желудок. Из средней кишки – тонкая кишка, печень и поджелудочная железа. Из задней кишки – слепая кишка и червеобразный отросток, восходящая ободочная, нисходящая ободочная, сигмовидная ободочная и прямая кишка. 24. Источники и ход эмбрионального развития органов мужской половой системы. В развитии половых органов различают 2 стадии: 1) индифферентной закладки, 2) дифференцировки по мужскому или женскому типу. У зародыша человека на 4-5-ой неделе внутриутробного развития определяются индифферентные гонады, расположенные на вентральной поверхности мезонефроса в виде утолщенного валика целомического эпителия. В гонаде формируются половые тяжи, определяются первичные половые клетки, которые проникают в закладку с током крови или через энтодерму задней кишки из желточного мешка. На 5-ой неделе эмбриологического развития вдоль латерального края первичной почки и мезонефрального протока формируется парамезонефральный проток. Из мезонефральных протоков формируются выводящие протоки мужских половых органов. На 7-8-ой неделе эмбриогенеза наступает дифференцировка индифферентной гонады по мужскому или женскому типу. Дифференцировка мужских половых органов происходит под влиянием тестостерона, который вырабатывают интерстициальные клетки (Лейдига). Они расположены в мезенхиме между половыми тяжами яичка. Интерстициальные клетки начинают функционировать на 3-ем месяце эмбриогенеза. Признаком дифференцировки гонады по мужскому типу является начало формирования белочной оболочки, а также редукция парамезонефральных протоков. Половые тяжи превращаются в извитые и прямые семенные канальцы, а из канальцев среднего отдела мезонефроса (первичной почки) развиваются канальцы сети и выносящие канальцы яичка. Краниальные канальцы первичной почки превращаются в привесок придатка яичка (appendix epididymidis), а каудально расположенные канальцы преобразуются в придаток привеска яичка (paradidymis). Наружные половые органы развиваются из индифферентных закладок. На 3-ем месяце эмбриогенеза кпереди от клоачной перепонки из мезенхимы образуется половой бугорок. От полового бугорка к анальному отверстию проходит уретральная борозда, которая ограничена половыми складками. По обеим сторонам от полового бугорка и мочеполовых складок формируются половые валики. После половой дифференцировки индифферентной гонады у зародышей мужского пола половой бугорок превращается в пещеристые тела полового члена. Половые складки и уретральная щель образуют мужскую уретру и губчатое тело полового члена. Половые валики формируют мошонку. Место срастания соответствует шву мошонки. 25. Источники и ход эмбрионального развития органов женской половой системы. В развитии половых органов различают 2 стадии: 1) индифферентной закладки, 2) дифференцировки по мужскому или женскому типу. У зародыша человека на 4-5-ой неделе внутриутробного развития определяются индифферентные гонады, расположенные на вентральной поверхности мезонефроса в виде утолщенного валика целомического эпителия. В гонаде формируются половые тяжи, определяются первичные половые клетки, которые проникают в закладку с током крови или через энтодерму задней кишки из желточного мешка. На 5-ой неделе эмбриологического развития вдоль латерального края первичной почки и мезонефрального протока формируется парамезонефральный проток. Из парамезонефральных протоков развиваются внутренние женские половые органы. На 7-8-ой неделе эмбриогенеза наступает дифференцировка индифферентной гонады по мужскому или женскому типу. Признаками дифференцировки гонады по женскому типу является отсутствие выраженной белочной оболочки, сужение просвета мезонефрального протока, первичные половые клетки разбросаны по всей мезенхимной строме. Часть первичных половых клеток растет активнее и они превращаются в фолликулы яичника (300-500). В дальнейшем образуется корковое и мозговое вещество яичника. Яичники смещаются вместе с маточными трубами в полость малого таза (опускание яичника). Причем маточные трубы из вертикального положения приобретают горизонтальное. Из краниального отдела парамезонефральных протоков развиваются моточные трубы, а из их дистальных сросшихся частей – матка и проксимальный отдел влагалища. Из мочеполового синуса формируется дистальный отдел влагалища и его преддверие. Канальцы мезонефроса и мезонефральный проток превращаются в придатки яичника (epoophoron, paraophoron). Наружные половые органы развиваются из индифферентных закладок. На 3-ем месяце эмбриогенеза кпереди от клоачной перепонки из мезенхимы образуется половой бугорок. От полового бугорка к анальному отверстию проходит уретральная борозда, которая ограничена половыми складками. По обеим сторонам от полового бугорка и мочеполовых складок формируются половые валики. У зародышей женского пола половой бугорок превращается в клитор. Половые складки превращаются в малые половые губы. Дистальная часть уретральной щели становится преддверием влагалища. Половые валики преобразуются в большие половые губы. 26. Источники и ход эмбрионального развития органов мочевыделительной системы. К мочевым органам относятся почки, мочеточники, мочевой пузырь и мочеиспускательный канал. Среди них почки являются мочеобразующими органами, а остальные составляют мочевыводящие пути. В течение эмбрионального периода закладываются последовательно три парных выделительных органа: передняя почка, или предпочка, первичная почка и постоянная, или окончательная, почка. Предпочка образуется из передних 8—10 сегментных ножек мезодермы. У зародыша человека предпочка не функционирует в качестве мочеотделительного органа и вскоре после закладки подвергается атрофии. Первичная почка формируется из большого числа сегментных ножек, расположенных в области туловища зародыша. Сегментные ножки отшнуровываются от сомитов и спланхнотома и превращаются в канальцы первичной почки. Возникший при развитии предпочки мезонефральный проток открывается в заднюю кишку. Канальцы растут по направлению к мезонефральному протоку, образующемуся при развитии предпочки, и вступают с ним в сообщение. Навстречу им от аорты отходят сосуды, распадающиеся на капиллярные клубочки. Канальцы своим слепым концом обрастают эти клубочки, образуя их капсулы. Капиллярные клубочки и капсулы вместе формируют почечные тельца. Окончательная почка закладывается у зародыша на 2-м месяце, но развитие ее заканчивается лишь после рождения ребенка. Эта почка образуется из двух источников — мезонефрального протока и нефрогенной ткани, представляющей собой не разделенные на сегментные ножки участки мезодермы в каудальной части зародыша. Мезонефральный проток дает начало мочеточнику, почечной лоханке, почечным чашечкам, сосочковым каналам и собирательным трубкам. Образовавшись, окончательная почка начинает быстро расти и с 3-го месяца оказывается лежащей выше первичной почки, которая во второй половине беременности атрофируется. 27. Источники и ход эмбрионального развития органов дыхательной системы. Дыхательная система — это совокупность органов, обеспечивающих в организме внешнее дыхание, а также ряд важных недыхательных функций. В состав дыхательной системы входят различные органы, выполняющие воздухопроводящую и дыхательную (газообменную) функции: полость носа, носоглотка, гортань, трахея, внелегочные бронхи и легкие. Носовая полость развивается из ротовой бухты. Гортань, трахея и легкие развиваются из одного общего зачатка, который появляется на 3—4-й неделе путем выпячивания вентральной стенки передней кишки. Гортань и трахея закладываются на 3-й неделе из верхней части непарного мешковидного выпячивания вентральной стенки передней кишки. В нижней части этот непарный зачаток делится по средней линии на два мешка, дающих зачатки правого и левого легкого. На 8-й неделе появляются зачатки бронхов в виде коротких ровных трубочек, а на 10—12-й неделе стенки их становятся складчатыми, выстланными цилиндрическими эпителиоцитами (формируется древовидно разветвленная система бронхов — бронхиальное дерево). На этой стадии развития легкие напоминают железу (железистая стадия). На 5—6-м месяце эмбриогенеза происходит развитие конечных (терминальных) и респираторных бронхиол, а также альвеолярных ходов, окруженных сетью кровеносных капилляров и подрастающими нервными волокнами (канальцевая стадия). Из мезенхимы, окружающей растущее бронхиальное дерево, дифференцируются гладкая мышечная ткань, хрящевая ткань, волокнистая соединительная ткань бронхов, эластические, коллагеновые элементы альвеол, а также прослойки соединительной ткани, прорастающие между дольками легкого. С конца 6-го — начала 7-го месяца и до рождения дифференцируется часть альвеол и выстилающие их альвеолоциты 1-го и 2-го типов (альвеолярная стадия). 28. Источники и ход эмбрионального развития органов зрения и слуха. Орган слуха по строению относится к вторичночувствующим органам (дендриты чувствительных нейронов подходят к сенсоэпителиальным клеткам). Развитие. Орган слуха имеет 3 источника развития: Эктодерма. Из неё развивается эпителий, сенсорные и поддерживающие клетки. Мезенхима. Она даёт начало соединительной ткани и сосудистым элементам. Нервная трубка. Это источник развития слухового и вестибулярного нервов. На 3-ей неделе эмбрионального развития на уровне ромбовидного мозга эктодерма симметрично утолщается, и образуются слуховые плакоды. Она инвагинируют и формируются слуховые ямки, которые вскоре превращаются в слуховые пузырьки. Слуховые пузырьки покрыты изнутри многорядным эпителием, который секретирует в просвет пузырька эндолимфу. С медиальной стороны к каждому слуховому пузырьку прилежит зачаток слухового ганглия, который развивается в ганглий преддверия и улитки. Одновременно с этим развиваются перилимфатические полости. Окружающая мезенхима даёт начало соединительной ткани и хрящевой бластеме, которая позже преобразуется в костную ткань. В слуховых пузырьках обособляются маточка и полукружные каналы, а также – мешочек с зачатком улитки. Улиточный канал отделяется от мешочка и развивается в виде трубки, которая постепенно вдаётся в завитки костной улитки. К концу 8-ой недели внутриутробного развития есть все разновидности сенсорных и поддерживающих клеток органов слуха и равновесия. Эпителий базальной стенки перепончатого канала даёт развитие спиральному органу. Постепенно формируются контакты между сенсорными клетками и дендритами чувствительных нейронов 8-ой пары ЧМН (n.vestibulocochlearis). Глаз развивается из нескольких источников. Из мозгового пузыря образуются 2 выпячивания — глазные пузырьки. Передняя стенка глазных пузырьков впячивается, в результате чего из каждого глазного пузырька образуется глазной бокал, связанный с нервной трубкой при помощи полого стебелька и состоящий из 2 стенок: наружной и внутренней. Из наружной стенки развивается пигментный слой сетчатки, из внутренней — нейронный слой сетчатки. Из краев глазного бокала развиваются мышца, суживающая зрачок, и мышца, расширяющая зрачок. Белочная и сосудистая оболочки, радужка, цилиарное тело и соединительнотканная основа роговицы глаза развиваются из мезенхимы; передний эпителий роговицы глаза и хрусталик — из кожной эктодермы. Развитие хрусталика происходит следующим образом. В то время, когда образуется глазной бокал, кожная эктодерма, расположенная напротив бокала, утолщается и впячивается в бокал. Это впячивание отделяется от эктодермы и в процессе развития превращается в хрусталик. Стекловидное тело развивается за счет мезенхимы с участием кровеносных сосудов. 29. Источники и ход эмбрионального развития органов эндокринной системы. Эндокринная система — совокупность структур: органов, частей органов, отдельных клеток, секретирующих в кровь и лимфу гормоны. В состав эндокринной системы входят специализированные эндокринные железы, или железы внутренней секреции, лишенные выводных протоков, но обильно снабженные сосудами микроциркуляторного русла, в которые выделяются продукты секреции этих желез. Источником развития эндокринных желёз являются разные зародышевые листки: 1. из энтодермы развиваются щитовидная, паращитовидная железы, тимус, панкреатические островки поджелудочной железы, одиночные эндокриноциты пищеварительного тракта и воздухоносных путей; 2. из эктодермы и нейроэктодермы - гипоталамус, гипофиз, мозговое вещество надпочечников, кальцитониноциты щитовидной железы; 3. из мезодермы и мезенхимы - корковое вещество надпочечников, половые железы, секреторные кардиомиоциты, юкстагломерулярные клетки почек. Зачаток щитовидной железы появляется у эмбриона человека в конце 4-й недели из эндодермы. На 7-й неделе происходит соединение боковых долей с перешейком. На 2-м месяце внутриутробного развития формируется тиреоидная ткань. 30. Источники и ход эмбрионального развития органов кроветворения и иммунной защиты. ИСТОЧНИКОМ РАЗВИТИЯ ОРГАНОВ КРОВЕТВОРЕНИЯ является мезенхима, за исключением тимуса, который развивается из эпителия III пары жаберных карманов. У человека процесс кроветворения начинается в конце 2-й — начале 3-й недели развития эмбриона. Костный мозг у человека появляется впервые на 2-м месяце внутриутробного периода в ключице эмбриона, затем на 3-4 -м месяце он образуется в развивающихся плоских костях, а также в трубчатых костях конечностей — лопатках, тазовых костях, затылочной кости, ребрах, грудине, костях основания черепа и позвонках, а в начале 4-го месяца развивается также в трубчатых костях конечностей. До 11-й недели это остеобластический костный мозг, который выполняет остеогенную функцию. В данный период костный мозг накапливает стволовые клетки, а клетки стромы с остеогенными потенциями создают микросреду, необходимую для дифференцировки стволовых кроветворных клеток. У 12—14-недельного эмбриона человека происходят развитие и дифференцировка вокруг кровеносных сосудов гемопоэтических клеток. У 20—28-недельного плода человека в связи с интенсивным разрастанием костного мозга отмечается усиленная резорбция костных перекладин остеокластами, в результате чего образуется костномозговой канал, а красный костный мозг получает возможность расти в направлении эпифизов. К этому времени костный мозг начинает функционировать как основной кроветворный орган, причем большая часть образующихся в нем клеток относится к эритроидному ряду гемопоэза. Уровень иммунной реактивности развивающихся зародышей значительно уступает половозрелым особям. Тем не менее, начальные этапы становления T- и B-звеньев иммунной системы проявляются очень рано. Тимус -- наиболее ранний орган иммунной системы, возникающий в процессе зародышевого развития. Формируется из эндодермы 3-го и 4-го глоточных карманов и эктодермы 3-й и 4-й жаберных щелей. Развивающийся тимус характеризуется интенсивной клеточной пролиферацией и увеличением массы органа. 31. Основные этапы развития эмбриологии. Вклад отечественных и зарубежных учёных в развитие эмбриологии. (К.Ф. Вольф, Х.И. Пандер, К.Э. Бэр, А. О. Ковалевский, А. Г. Кнорре, О. В. Волкова, А. И. Никитин и др.). Эбриология (от эмбрион и логос — слово, учение), наука о предзародышевом развитии (образование половых клеток), оплодотворении, зародышевом и личиночном развитии организма. Выделяют эмбриологию животных и человека и эмбриологию растений. Различают общую, сравнительную, экспериментальную и экологическую эмбриологию. Основоположники эмбриологии — Гиппократ и Аристотель, а в новое время — К. Ф. Вольф и К. М. Бэр; А. О. Ковалевский и И. И. Мечников заложили основы эволюционной эмбриологии. Решительный поворот в эмбриологии был осуществлен в 1759 г. петербургским академиком Каспаром Фридрихом Вольфом (1734–1794). В этом году Вольф представил свою диссертацию под названием «Теория зарождения». В те времена господствовало мнение физиолога и анатома А.Галлера о том, что трубчатые и мешкоподобные структуры зародыша (например, его кишечник) с самого начала имеют такую же форму, но это трудно заметить из-за тонкости стенок и их плотного слипания. Позже происходит их простое раздувание. Такое толкование строго соответствовало преформационной теории. Вольф установил совершенно иное. Кишечник, а также зачаток нервной системы сначала представляют собой пласты, которые лишь позже скручиваются в трубки. В ходе развития образуются новые формы. По сути дела Вольф открыл формообразование и тем самым дал первый позитивный и неопровержимый аргумент в пользу эпигенеза. Судьба этих, казалось бы, столь ясных работ была трудной. Под давлением господствующих авторитетов выводы Вольфа отвергались, и его работы были на некоторое время забыты. Дальнейший прогресс в эмбриологии позвоночных связан с именами М.Ратке (1793–1860), X.Пандера (1794–1865) и К.Бэра (1792–1876). Пандер в 1817г. впервые описал зародышевые листки. Он нашел, что зародыш цыпленка на определенной стадии состоит из трех пластов: наружного – серозного, самого глубокого – слизистого и промежуточного – кровяного. Гипотеза Дарвина оказалась мощным стимулом к эмбриологическим исследованиям. На основе эволюционной теории ученые разных стран за считанные годы выяснили развитие обширных, ранее совершенно не изученных групп организмов. Среди них первыми были русские эмбриологи А.О.Ковалевский (1840–1901) и И.И.Мечников (1845–1916). Особое значение имели работы Ковалевского по развитию ланцетника и асцидий, в которых были продемонстрированы сходные черты в развитии позвоночных и беспозвоночных животных. Эти исследования способствовали укреплению взглядов на эволюцию как на сквозной монофилетический процесс. А.О.Ковалевский – один из основоположников теории зародышевых листков. III. ОБЩАЯ ГИСТОЛОГИЯ 1. Возникновение тканей на основе дифференциации клеток эмбриональных зачатков. Механизмы гистогенезов. Дифференцировка зародышевых листков и мезенхимы начинается в конце 2-й недели. Одна часть клеток преобразуется в зачатки тканей и органов зародыша, другая — во внезародышевые органы. Дифференцировка зародышевых листков и мезенхимы, приводящая к появлению тканевых и органных зачатков, происходит неодновременно, но взаимосвязанно. Формирование тканевых зачатков происходит на основе процессов детерминации и коммитирования. Дифференциация— это стойкое структурно-функциональное преобразование клеток в различные специализированные клетки. Дифференцировка клеток биохимически связана с синтезом специфических белков, а цитологически — с образованием специальных органелл и включений. Детерминация— это процесс определения пути, направления, программы развития материала эмбриональных зачатков с образованием специализированных тканей. Ткани - это исторически (филогенетически) сложившиеся системы клеток и неклеточных структур, обладающих общностью строения, в ряде случаев - общностью происхождения, и специализированные на выполнении определенных функций. Ткани развиваются путем гистогенеза. Гистогенез - единый комплекс координированных во времени и пространстве процессов пролиферации, дифференцировки, детерминации, интеграции и функциональной адаптации клеточных систем. Развитие организма начинается с одноклеточной стадии — зиготы. В ходе дробления возникают бластомеры, но совокупность бластомеров – это еще не ткань. Бластомеры на начальных этапах дробления еще не детерминированы (они тотипотентны). Если отделить их один от другого, - каждый может дать начало полноценному самостоятельному организму – механизм возникновения монозиготных близнецов. Постепенно на следующих стадиях происходит ограничение потенций. В основе его лежат процессы, связанные с блокированием отдельных компонентов генома клеток и детерминацией. Эмбриональный гистогенез — процесс возникновения специализированных тканей из малодифференцированного клеточного материала эмбриональных зачатков, происходящий в течение эмбрионального развития организма. Эмбриональные зачатки — источники развития тканей и органов в онтогенезе, представленные группами малодифференцированных клеток. Гистогенез сопровождается размножением и ростом клеток, их перемещением — миграцией, дифференцировкой клеток и их производных, межклеточными и межтканевыми взаимодействиями — корреляциями, отмиранием клеток. На разных этапах индивидуального развития могут иметь преимущественное значение те или иные из перечисленных компонентов. 2. Ткани, как один из уровней организации живого. Определение. Принципы классификации тканей. Вклад отечественных и зарубежных ученых в учение о тканях. Ткань — это система клеток и неклеточных структур, обладающая общностью строения, а иногда и происхождения, и специализированная на выполнении определенных функций. Характеристика структурных компонентов ткани: Клетки — основные, функционально ведущие компоненты тканей. Все ткани состоят из нескольких типов клеток. Клеточная популяция — это совокупность клеток данного типа. Клеточный дифферон, или гистогенетический ряд, — это совокупность клеток данного типа (данной популяция), находящихся на различных этапах дифференцировки. Производные клеток: 1) симпласты (слияние отдельных клеток, например мышечное волокно); 2) синцитий (несколько клеток, соединенных между собой отростками, например сперматогеиный эпителий извитых канальцев семенника); 3) постклеточные образования (эритроциты, тромбоциты). Межклеточное вещество — также продукт деятельности определенных клеток. Межклеточное вещество состоит из: 1) аморфного вещества; 2) волокон (коллагеновых, ретикулярных, эластических). Классификации тканей: 1) эпителиальные ткани; 2) соединительные ткани (ткани внутренней среды, опорно-трофические ткани); 3) мышечные ткани; 4) нервная ткань. Московская школа гистологов была создана одним из крупных представителей материалистического направления в естествознании 19века – А.И. Бабухиным. Большое внимание уделялось вопросам гистогенеза различных тканей. А.А. Заварзин считал основной задачей гистологии – выяснение общих закономерностей филогенетической дифференцировки разновидностей специализированных клеток в пределах каждой ткани при сохранении ограниченного числа морфофункциональных типов тканей. Н. Г. Хлопин сделала обобщение в области изучения эволюционного развития тканей. Знание нормальной структуры клеток, тканей и органов является необходимым условием для понимания механизмов изменений в низ в патологических условиях. Поэтому гистология тесно связана с патологической анатомией и многими клиническими дисциплинами. 3. Стволовые клетки и их свойства. Детерминация и дифференциация клеток в ряду последовательных делений, коммитирование потенций. Стволовые - это клетки с низкой степенью дифференцировки, которые могут неограниченное время делиться и таким образом поддерживать свою популяцию, превращаться в разные типы клеток под действием различных стимулов. Основным источником стволовых клеток во взрослом организме является костный мозг. Стволовые клетки способны выходить из костного мозга в кровь, а затем - в органы и ткани. Поэтому, в каждом органе и ткани в настоящее время обнаружены стволовые клетки. В органах и тканях стволовые клетки могут так и оставаться стволовыми, а могут подвергаться дальнейшей дифференцировке, то есть превращаться в зрелые специфические клетки, например, мышечные, эпителиальные, соединительнотканные, крови и т.д. Дифференцировку стволовых клеток индуцируют факторы роста, колониестимулирующие факторы, гормоны, цитокины, их микроокружение и др. Массовый выход стволовых клеток из костного мозга называется мобилизацией. Одними из изученных факторов, способствующих мобилизации, являются колониестимулирующий фактор гранулоцитов-моноцитов, эритропоэтин. Стволовые клетки обеспечивают рост, физиологическое обновление и репаративную регенерацию органов и тканей. Возможно, стволовые клетки участвуют в возникновении, росте и метастазировании злокачественных опухолей. Классификация: Недифференцированные (эмбриональные). Они появляются у эмбриона и сохраняются во взрослом организме. Гематопоэтические. Мезенхимальные - представляют собой разновидность клеток стромы костного мозга. Эндотелиальные. Тканевые - располагаются в органах и тканях. Возможно, что тканевые стволовые клетки являются потомками гематопоэтических, мезенхимальных или эндотелиальных стволовых клеток. Детерминация – это процесс определения дальнейшего пути развития материала эмбриональных зачатков с образование специфических тканей (на основе блокирования отдельных генов). Понятие «коммитирование» тесно связано с клеточным делением (т.н. коммитирующий митоз). Коммитирование – это ограничение возможных путей развития вследствие детерминации. Коммитирование совершается ступенчато. Сначала соответствующие преобразования генома касаются крупных его участков. Затем все более детализируются, поэтому вначале детерминируются наиболее общие свойства клеток, а затем и более частные. 4. Пределы изменчивости тканей. Современные представления о дифферонах, ”тканевых мозаиках”. Значение гистологии для медицины. Регенерация и пределы изменчивости тканей. Восстановительная способность тканей - это их способность к рененерации. Регенерация - биологический процесс, обеспечивающий восстановление погибших или утраченных частей (элементов). Существует регенерация: 1) Физиологическая - восстановление тканей после естественного изнашивания в процессе жизнедеятельности; 2) репаративная- восстановление после повреждения или утраты. Однако не во всех тканях они в равной степени представлены, а в некоторых тканях могут отсутствовать (нервная ткань, сердечная мышечная ткань). В связи с этим все ткани подразделяют на 3 группы: 1) ткани с обновляющимися клетками; 2) ткани с лабильными клетками; 3) ткани со стационарными клетками. Степень физиологической и репаративной регенерации у них разная. Изменчивость тканей - это их способность изменять свои свойства в зависимости от возраста и условий окружающей среды. Возрастные изменения связаны с уменьшением численности клеток, снижением в них обменных процессов, что приводит к дистрофическим изменениям клеточных и неклеточных структур тканей. Изменения под воздействием средовых факторов отражают адаптацию тканей к сложившимся условиям существования, что чаще всего проявляется в компенсаторном усилении митотической активности и метаболических процессов, приводящих к гипертрофии и гиперплазии клеточных элементов. Клеточный дифферон —это совокупность клеточных форм, составляющих ту или иную линию дифференцировки от стволовой до терминально дифференцированной клетки.Начальной клеткой клеточного дифферона является стволовая клетка. Следующую стадию гистологического ряда образуют полустволовые (коммитированные) клетки, которые в отличие от стволовых клеток могут дифференцироваться только в каком-то одном направлении. Третьей и самой многочисленной частью дифферона являются дифференцированные, функционально активные клетки. Наконец, четвертым компонентом являются старые, функционально неактивные клетки и постклеточные структуры. Важная задача общей Гистология — выяснение потенций развития, присущих каждому типу дифференцированных клеток, и механизмов, регулирующих сохранение постоянства дифференцировки и ее изменения. В каждой ткани различают несколько устойчивых типов клеточной дифференцировки, например фибробласты, образующие основное вещество соединительной ткани, и эритроидные клетки, образующие и несущие дыхательные пигменты. Каждый тип дифференцировки достигается в ходе многоэтапного процесса развития ткани — гистогенеза. В клетках, выполняющих специализированные функции, реализуется лишь небольшая часть возможностей, предусмотренных генетической программой организма. Остальная, не реализуемая в дифференцированных клетках часть генетической информации сохраняется в них, но находится в неактивном, или репрессированном, состоянии. 5. Закономерности возникновения и эволюции тканей. Теории параллелизма А. А. Заварзина и дивергентной эволюции тканей Н. Г. Хлопина, их синтез на современном уровне развития (А. А. Браун, В. П. Михайлов). В ходе эволюции происходило возникновение, развитие и усложнение строения различных тканей. Ход эволюции тканей наиболее полно объясняют следующие теории. Теория параллельных рядов. А.А. Заварзин разработал теорию эволюции тканей, которая называется теорией параллельных рядов тканевой эволюции, или теорией параллелизма. Суть этой теории заключается в том, что в ходе эволюции в разных ветвях филогенетического дерева самостоятельно, независимо, параллельно возникали одинаково построенные ткани, выполняющие сходные функции. Например, соединительная ткань ланцетника и млекопитающих выполняет одинаковые функции и поэтому имеет общие черты строения. Теория параллельных рядов хорошо раскрывает причины эволюции тканей, а также возможности их адаптации. Теория дивергентного развития тканей. Н.Г. Хлопин предложил собственную оригинальную теорию эволюции тканей, которая называется теорией дивергентного развития тканей. Согласно этой теории, ткани в эволюции и онтогенезе развиваются дивергентно, то есть возникают из уже существующих тканей путем расхождения признаков, что ведет ко все возрастающему разнообразию тканей. Эта теория показывает, как в ходе дивергенции из одного эмбрионального зачатка образуются ткани, постепенно приобретающие все более выраженные различия в строении и функциях. Например, развивающиеся из кожной эктодермы эпидермис и многослойный плоский эпителий имеют больше сходств, чем различий, тогда как имеющие общий с ними источник развития эпителий аденогипофиза, эмаль зуба и др. разительно от них отличаются. Единая концепция эволюционного развития тканей. Теории А.А. Заварзина и Н.Г. Хлопина органично дополняют друг друга. Поэтому советские гистологии А.А. Браун, В.П. Михайлов объединили их в единую теорию эволюции тканей, которая утверждает, что сходные тканевые структуры в различных ветвях филогенетического дерева возникли параллельно в ходе дивергентного развития. 6. Восстановительные способности тканей, типы физиологической регенерации в обновляющихся, растущих и стационарных клеточных популяциях. Репаративная регенерация. Регенерация — восстановление клеток, направленное на поддержание функциональной активности данной системы. В регенерации различают такие понятия, как форма регенерации, уровень регенерации, способ регенерации. Формы регенерации: физиологическая регенерация — восстановление клеток ткани после их естественной гибели (например, кроветворение); репаративная регенерация — восстановление тканей и органов после их повреждения (травмы, воспаления, хирургического воздействия и так далее). Уровни регенерации — соответствуют уровням организации живой материи: клеточный (внутриклеточный); тканевой; органный. Эмбриональная (быстро делящаяся клеточная популяция) – все клетки популяции активно делятся, специализированные элементы отсутствуют. Стабильная клеточная популяция – долгоживущие, активно функционирующие клетки, которые вследствие крайней специализации утратили способность к делению. Например, нейроны, кардиомиоциты. Растущая (лабильная) клеточная популяция – специализированные клетки которой способны делиться в определённых условиях. Например, эпителии почки, печени. Репаративная (от лат. reparatio — восстановление) регенерация наступает после повреждения ткани или органа. Она очень разнообразна по факторам, вызывающим повреждения, по объемам повреждения, по способам восстановления. Механическая травма, например оперативное вмешательство, действие ядовитых веществ, ожоги, обморожения, лучевые воздействия, голодание, другие болезнетворные агенты,— все это повреждающие факторы. Наиболее широко изучена регенерация после механической травмы. Способность некоторых животных, таких, как гидра, планария, некоторые кольчатые черви, морские звезды, асцидия и др., восстанавливать утраченные органы и части организма издавна изумляла ученых. Ч. Дарвин, например, считал удивительными способность улитки воспроизводить голову и способность саламандры восстанавливать глаза, хвост и ноги именно в тех местах, где они отрезаны. Объем повреждения и последующее восстановление бывают весьма различными. Крайним вариантом является восстановление целого организма из отдельной малой его части, фактически из группы соматических клеток. IV. ЭПИТЕЛИАЛЬНЫЕ ТКАНИ 1. Эпителиальные ткани, источники их развития, общая морфофункциональная характеристика. Классификации. Значение работ Н. Г. Хлопина, А. А. Заварзина, Ф. М. Лазаренко для изучения эпителиальных тканей. Эпителиальные ткани — это совокупность дифферонов полярно дифференцированных клеток, тесно расположенных в виде пласта на базальной мембране, на границе с внешней или внутренней средой, а также образующих большинство желез организма. Эпителиальные ткани представляют собой пласт клеток, лежащих на базальной мембране, — пограничное расположение. Эпителиальные ткани образованы одним видом тканевых элементов — клетками. Эпителиоциты всех эпителиальных тканей содержат в цитоплазме особый вид промежуточных филамент, которые называются кератиновыми филаментами и состоят из цитокератинов различного типа. В эпителиях отсутствуют собственные сосуды, их питание идет через базальную мембрану путем диффузии веществ из сосудов соединительной ткани (единственное исключение — эпителий сосудистой полоски внутреннего уха). Эпителий, как правило, хорошо иннервируется. Для эпителиальных тканей характерны 5 основных функций — пограничная, барьерно-защитная, секреторная, экскреторная (выделительная), всасывательная (резорбтивная). Источники развития эпителиальных тканей. Эпителии развиваются из всех трех зародышевых листков, начиная с 3—4-й недели эмбрионального развития человека. В зависимости от эмбрионального источника различают эпителии эктодермального, мезодермального и энтодермального происхождения. Родственные виды эпителиев, развивающиеся из одного зародышевого листка, в условиях патологии могут подвергаться метаплазии, т.е. переходить из одного вида в другой. Классификация тканей: А) Функциональная классификация: 1) покровный эпителий; 2) железистый (секреторный) эпителий, 3) сенсорный (чувствительный) эпителий. Б) Морфологическая классификация: I. Однослойный эпителий 1. Многорядный 2. Однорядный а) плоский 6) кубический в) призматический II. Многослойный эпителий 1. Плоский; а) неороговевающий 6) ороговевающий 2. Кубический 3. Призматический 4. Переходный В) Генетическая классификация (по источникам развития) (по Н.Г. Хлопину). 1. Эпителий эктодермального типа. Развивается из кожной эктодермы. 2. Эпителий энтодермального типа. Источником развития для этого эпителия служит кишечная энтодерма. 3. Эпителий целонефродермального типа. Развивается из мезодермы: листков спланхнотома (мезотелий, эпителий коры надпочечника), нефротома (эпителий канальцев нефрона). 4. Эпителий эпендимоглиального типа развивается из нейроэктодермы (эпендима, задний эпителий роговицы, периневральный эпителий). 5. Эпителий ангиодермального типа (эндотелий). Развивается из мезенхимы. А.А. Заварзин на основе глубокого сравнительно-гистологического изучения нервной системы сформулировал принцип параллелизма тканевых структур, переработанный позднее в теорию тканевой эволюции. Он сделал вывод, что все животные имеют общий принцип тканевой организации и состоят из 4 тканевых систем. Это связано с тем, что всякий организм находится в одинаковых условиях взаимодействия с окружающей средой и выполняет 4 наиболее общие функции — защитную, внутреннего обмена и постоянства внутренней среды, движения, реактивности. А.А. Заварзин обосновал морфофункциональную классификацию тканей. Теория А.А. Заварзина называется теорией параллельных рядов тканевой эволюции. Хлопин разработана теория дивергентного развития тканей. Согласно этой теории, эволюционное развитие тканей происходит принципиально так же, как и организмов, подчиняясь тем же основным закономерностям, что и целые организмы. Как известно, организмы развиваются дивергентно, т.е. расхождением признаков, благодаря чему и возникает многообразие форм. Дивергентная эволюция тканей имеет, однако, свою специфику, в силу чего многообразие тканей ограничено известными пределами признаков, свойственных 4 основным системам тканей. Основные научные достижения Ф. М. Лазаренко касаются области гистологии соединительной ткани и эпителиальносоединительнотканных взаимоотношений. В тридцатые годы Ф. М. Лазаренко, работавший в Оренбурге, разработал оригинальный метод культивирования тканей и органов в организме. Под руководством Ф. М. Лазаренко сформировалась большая школа гистологов-экспериментаторов, внесших достойный вклад в развитие биологии, медицины и ветеринарии. 2. Строение эпителиальных тканей. Строение и роль базальной мембраны. Физиологическая и репаративная регенерация эпителиев. Эпителий, выстилающий все поверхности в организме, имеет вид пласта, тогда как в печени, поджелудочной, щитовидной, слюнной и других железах он представляет собой скопление клеток. Клетки лежат плотно друг к другу, образуя сплошной пласт. Гетерополярность - апикальные (верхушка) и базальные части клеток отличаются по строению и по функции; а в многослойном эпителии - отличие в строении и функции слоев. Состоит только из клеток, межклеточное вещество практически отсутствует (десмосомы). Эпителий всегда располагается на базальной мембране (углеводнобелковолипидный комплекс с тончайшими фибриллами) и им отграничивается от подлежащей рыхлой соединительной ткани. Эпителий участвует в выделении секрета. Характерна повышенная регенераторная способность, обусловленная пограничностью. Не имеет собственных кровеносных сосудов, питается диффузно через базальную мембрану, за счет сосудов подлежащей рыхлой соединитткани. Хорошо иннервирован (много нервных окончаний). Эпителии располагаются на базалъных мембранах (пластинках), которые образуются в результате деятельности как клеток эпителия, так и подлежащей соединительной ткани. Базальная мембрана имеет толщину около 1 мкм и состоит из подэпителиальной электронно-прозрачной светлой пластинки (lamina lucida) толщиной 20—40 нм и темной пластинки (lamina densa) толщиной 20— 60 нм (рис. 53). Светлая пластинка Включает аморфное вещество, относительно бедное белками, но богатое ионами кальция. Темная пластинка имеет богатый белками аморфный матрикс, в который впаяны фибриллярные структуры (коллаген IV типа), обеспечивающие механическую прочность мембраны. В ее аморфном веществе содержатся сложные белки — гликопротеины, протеогли- каны и углеводы (полисахариды) — гликозаминогликаны. Гликопротеины — фибронектин и ламинин — выполняют роль адгезивного субстрата, с помощью которого к мембране прикрепляются эпителиоциты. Базальная мембрана выполняет ряд функций: механическую (прикрепительную), трофическую и барьерную (избирательный транспорт веществ), морфогенетическую (организующую при регенерации) и ограничивающую возможность инвазивного роста эпителия. Физиологическая регенерация – восстановление организмом утраченных или поврежденных органов или тканей. Репаративная регенерация –восстановление какой – либо ткани в патологических условиях. 3. Общая морфофункциональная характеристика, классификация многослойных эпителиев. Многослойные эпителии - Эпителиальный пласт представлен несколькими слоями эпителиоцитов - С базальной мембраной связан только базальный (самый глубокий) слой эпителиоцитов, в его составе много камбиальныхклеток Клетки вышележащих слоев теряют связь с базальной мембраной (источником их трофического обеспечения) и утрачивают митотическую активность. - Своими базальными полюсами они связаны с апикальными полюсами нижележащих эпителиоцитов. - Клетки поверхностного слоя постоянно отторгаются в окружающее пространство и заменяются новыми, пришедшими из нижележащих слоев. - Среди этих эпителиев выделяют: многослойный плоский неороговевающий, многослойный плоский ороговевающий, переходный, многослойный призматический. • Многослойный плоский неороговевающий (эпителий роговицы глаза) или частично ороговевающий (слизистые оболочек полости рта, пищевода, влагалища) - Характеризуется отсутствием или незначительностью (непостоянностью) процессов ороговения в эпителиоцитах - Клетки поверхностного слоя имеют уплощенную форму В этом эпителии выделяют 3 слоя клеток: базальный слой образован эпителиоцитами призматической формы, расположенных на базальной мембране. Среди этих клеток имеются стволовые, способные к делению. Поэтому его ещё называют камбиальным. шиповатый слой образован эпителиоцитами полигональной формы, имеющих отростки, которые вдаются между апикальной частью клеток базального слоя. Эти клетки располагаются несколькими слоями и имеют цитоплазматические отростки, которые напоминают шипы. слой плоских клеток, которые располагаются поверхностно и представляют отмирающие клетки. • Многослойный плоский ороговевающий эпителий Эпителиоциты (кератиноциты), потеряв связь с источником питания, ороговевают - Эпителиоциты поверхностного (рогового) слоя превращаются в плоские роговые чешуйки (корнеоциты – постклеточные формы) и слущиваются, по форме поверхностных клеток этот эпителий всегда плоский, 5 слоев(базальный, шиповатый, зернистый, блестящий, роговой). • Переходный эпителий — выстилает большую часть мочевыводящих путей ( лоханки, мочеточники, мочевой пузырь). При растяжении этих органов изменяется высота эпителиоцитов и количество клеточных рядов в пластах эпителия. Образован 3 слоями клеток: Базальный - образован мелкими круглыми (темными) клетками. Промежуточный - образован клетками полигональной формы. Поверхностный - образован крупными клетками, имеющими куполообразную и уплощённую форму, содержащими нередко 2 и 3 ядра. •Многослойный призматический - выстилает один из отделов прямой кишки. Состоит из нескольких слоев призматических эпителиоцитов. 4. Общая морфофункциональная характеристика, классификация однослойных эпителиев. Однослойные эпителии. - Все клетки эпителиального пласта связаны с базальной мембраной. - Апоптирующие и стареющие клетки постоянно заменяются новыми, образовавшимися в следствии митозов камбиальных эпителиоцитов - Среди этих эпителиев выделяют однорядные и многорядные • Однорядные эпителии - эпителиоциты, входящие в пласт, могут (не всенгда) иметь различное строение и выполнят разные функцииодинаковы - они одинковы по высоте и поэтому их ядра располагаются на одном уровне, т.е. в один ряд. Такие эпителии по форме эпителиоцитов делят на плоские, низкопризматические (кубические), высокопризматические (цилиндрические) - плоский эпителий составлен эпителиоцитами, у которых высота меньше их основания (например: эпителий кровеносных сосудов - эндотелий, эпителий серозных оболочек – мезотелий, эпителий легочных альвеол). -низкопризматический эпителий составлен эпителиоцитами, у которых высота равна их основанию (например: эпителий нефронов почек). - высокопризматический эпителий составлен эпителиоцитами, у которых высота больше их основанию (например: эпителий желудка, кишечника, канальцев нефронов почки). Однослойный многорядный (псевдомногслойный) – выстилает воздухоносные пути и некоторые отделы половой системы. В этом эпителии различают 4 вида клеток: 1) реснитчатые (мерцательные) клетки; 2) короткие и длинные вставочные клетки; 3) слизистые (бокаловидные) клетки; 4) базально - зернистые ( эндокринные) клетки; Вставочные клетки, вероятно, являются стволовыми, способными делиться и превращаться в реснитчатые и слизистые клетки. Вставочные клетки имеют форму клина и широкой частью прикрепляются к базальной мембране , а узкая вклинивается между реснитчатыми клетками , не достигая поверхности эпителия. Реснитчатые (мерцательные) клетки имеют форму клина, узкая часть клетки прикрепляется к базальной мембране, а широкая выходит на поверхность эпителия и имеет реснички. Слизистые клетки бокаловидной или овоидной формы, выделяют муцины на поверхность эпителия. Эндокринные (базально - зернистые) продуцируют биологически активные вещества ( гормоны ). Благодаря разному расположению вставочных и реснитчатых клеток в этом эпителии можно выделить три ряда ядер : нижний и средний ряды - ядра вставочных клеток , верхний ряд - ядра реснитчатых клеток. 5. Железистый эпителий. Особенности строения секреторных эпителиоцитов. Цитологическая характеристика эпителиоцитов, выделяющих секрет по голокриновому, апокриновому и мерокриновому типу. Железистый эпителий образован железистыми клетками - гландулоцитами, которые осуществляют синтез, а также выделение специфических секретов на поверхность кожи, слизистых оболочек и в полость внутренних органов (внешняя, экзокринная секреция) или в кровь и лимфу (внутренняя, эндокринная секреция). Гландулоциты (экзокриноциты) с внешней секрецией имеют секреторные включения в цитоплазме, развитую эндоплазматическую сеть, полярное расположение органелл и секреторных гранул. Железистый эпителий образует железы, которые делятся на 2 большие группы по выделению секрета: 1. Экзокринные внешней секреции, продуцируют секрет на поверхность эпителия. Экзокринные железы все без исключения имеют два отдела: а) концевой (секреторный) б) выводной проток 2. Эндокринные - внутренней секреции, не имеют выводных протоков и их продукты выделяются непосредственно в кровь или лимфу. Кроме этого все железы классифицируются по строению, типу секреци, а также по характеру секрета, расположению в эпителиальном пласте. По типу секреции (способ выделения секрета) железы делятся на: 1) мерокриновые; 2) апокриновые; 3) голокриновые. Мерокриновые - секрет выделяется без разрушения целостности клетки. Большинство желез в организме человека мерокриновые (слюнные, потовые). Апокриновые - секрет выделяется с отторжением апикальной части клетки. У человека - молочные железы и специфические потовые железы. Этот тип секреции еще называют макроапокриновым. Существует и микроапокриновый, когда разрушаются только верхушки микроворсинок . Голокриновые - это железы, клетки которых после накопления секрета полностью разрушаются, а ее остатки входят в состав секрета. У людей по голокриновому типу секретируют сальные железы. Название происходит от греческого слова hоlos - целый, krino - выделяю. 6. Железы, их классификация. Характеристика концевых отделов и выводных протоков экзокринных желез. Особенности строения эндокринных желез. Железистая эпителиальная ткань формирует железы — органы, состоящие из секреторных клеток, вырабатывающих и выделяющих специфические вещества различной химической природы. Вырабатываемые железами секреты имеют важное значение для процессов пищеварения, роста, развития, взаимодействия с внешней средой и других. Многие железы — самостоятельные, анатомически оформленные органы (например, поджелудочная железа, крупные слюнные железы, щитовидная железа), некоторые являются лишь частью органов (например, железы желудка). Железы подразделяются на две группы: железы внутренней секреции, или эндокринные, и железы внешней секреции, или экзокринные. И те и другие железы могут быть одноклеточными и многоклеточными. Экзокринные вырабатывают секреты, выделяющиеся на поверхность кожи, в полость органов. Они могут быть одноклеточными (бокаловидные клетки) и многоклеточными. Многоклеточные состоят из 2-х частей: секреторного отдела и концевых отделов. Концевые отделы образованы гландулоцитами, которые лежат на базальной мембране. По строению концевых отделов различают железы: разветвленные и неразветвленные, а также трубчатые, альвеолярные или смешанные. По строению выводных протоков различают железы: простые и сложные. Простые железы имеют неветвящийся выводной проток, сложные железы — ветвящийся. Эндокринные железы поставляют свой секрет (инкрет) во внутреннюю среду (кровь, лимфу, тканевую жидкость), а экзокринные – во внешнюю. Главным отличием эндокринных желез является то, что они не имеют выводных протоков и богато васкуляризированы. Инкрет содержит высокоактивные биологические вещества – гормоны, которые вместе с нервной системой участвуют в регулирующих функциях. Эндокринные железы или железы внутренней секреции (в дальнейшем сокращенно ЖВС) - это органы, состоящие из железистых специфических клеток, специализирующихся на образовании и выделении во внутреннюю среду организма особых биологически активных веществ - гормонов, участвующих в регуляции и интеграции функций организма. V. КРОВЬ, ГЕМОПОЭЗ, ИММУНОЦИТЫ 1. Понятие о системе крови. Кровь, как разновидность тканей внутренней среды. Функции крови, возрастные и половые особенности крови. Форменные элементы крови. Формула крови. Система крови – совокупность органов кроветворения, форменных элементов периферической крови, органов кроверазрушения и регуляторного аппарата. Понятие системы крови было введено в 1830-х гг. Х. Лангом. Кровь – это физиологическая система, которая включает в себя: · периферическую (циркулирующую и депонированную) кровь; · органы кроветворения; · органы кроверазрушения; · механизмы регуляции. Система крови включает в себя следующие компоненты: 1)кровь и лимфу; 2)органы кроветворения и иммунопоэза; 3)клетки крови, выселившиеся из крови в соединительную и эпителиальную ткани и способные вернуться (рециркулировать) снова в кровеносное русло (лимфоциты). Кровь как ткань обладает следующими особенностями: 1. все ее составные части образуются за пределами сосудистого русла; 2. межклеточное вещество ткани является жидким; 3. основная часть крови находится в постоянном движении. Функции крови: дыхательная: транспортирует кислород к тканям от легких и углекислый газ от тканей к легким. трофическая: переносит питательные вещества от стенки пищеварительного тракта к тканям. обменная: участвует в вводно-солевом обмене. экскреторная: переносит конечные продукты обмена от тканей к почкам. гомеостатическая: участвует в поддержании постоянства внутренней среды организма. регуляторная: переносит гормоны и другие биологически активные вещества, обеспечивая гуморальную регуляцию. терморегуляционная: кровь согревается в печени и мышцах и распределяет и перераспределяет тепло в организме. защитная: в крови имеются антитела; лейкоциты выполняют функцию фагоцитоза генетически чужеродных частиц; кровь способна свертываться, предотвращая кровопотерю. Возрастные особенности: Количество крови у взрослого человека составляет в среднем 7 % веса тела, у новорожденных - от 10 до 20 % веса тела, у грудных детей - от 9 до 13 %, у детей с 6 до 16 лет - 7 %. Плазма представляет собой жидкую часть крови после отделения всех форменных элементов. На ее долю у взрослых приходится 55-60 % общего объема крови, у новорожденных - меньше 50 % вследствие большого объема эритроцитов. Вязкость крови взрослого человека составляет 4-5, новорожденного - 10-11, ребенка первого месяца жизни - 6, затем наблюдается постепенное снижение вязкости. форменных элементов: эритроцитов (красных клеток крови), лейкоцитов (белых клеток крови), тромбоцитов(кровяных пластинок). Лейкоцитарная формула крови — это соотношение различных видов лейкоцитов, обычно выраженное в процентах. Х=А*В/Б*4000 Где а - общее количество эритроцитов, полученное при подсчете б - число малых квадратов в которых производился подсчет (80) в разведение крови (1:100, 1:200). 4000 - величина обратная объему жидкости на малым квадрате. 2. Эритроциты: размеры, форма, строение, функции, классификация по форме, размерам и степени зрелости. Особенности строения плазмолеммы эритроцита и его цитоскелета. Виды гемоглобина и связь с формой эритроцита. Ретикулоциты. Эритроциты, или красные кровяные тельца, человека и млекопитающих представляют собой безъядерные клетки, утратившие в процессе фило- и онтогенеза ядро и большинство органелл. Эритроциты являются высокодифференцированными постклеточными структурами, неспособными к делению. Размеры. Эритроцитов в нормальной крови также варьируют. Большинство эритроцитов (75 %) имеют диаметр около 7,5 мкм и называются нормоцитами. Остальная часть эритроцитов представлена микроцитами (~ 12,5 %) и макроцитами (~ 12,5 %). Микроциты имеют диаметр < 7,5 мкм, а макроциты >7,5 мкм. Изменение размеров эритроцитов встречается при заболеваниях крови и называется анизоцитозом. Форма и строение. Популяция эритроцитов неоднородна по форме и размерам. В нормальной крови человека основную массу (80—90 %) составляют эритроциты двояковогнутой формы — дискоциты. Кроме того, имеются планоциты (с плоской поверхностью) и стареющие формы эритроцитов — шиловидные эритроциты, или эхиноциты (~ 6 %), куполообразные, или стоматоциты (~ 1—3 %), и шаровидные, или сфероциты (~ 1 %). Химический состав. Плазмолемма. Плазмолемма эритроцита состоит из бислоя липидов и белков, представленных приблизительно в равных количествах, а также небольшого количества углеводов, формирующих гликокаликс. Большинство липидных молекул, содержащих холин (фосфатидилхолин, сфин-гомиелин), расположены во внешнем слое плазмолеммы, а липиды, несущие на конце аминогруппу (фосфатидилсерин, фосфатидилэтаноламин), лежат во внутреннем слое. Часть липидов (~ 5 %) наружного слоя соединены с молекулами олигосахаров и называются гликолипидами. Распространены мембранные гликопротеины — гликофорины. С ними связывают антигенные различия между группами крови человека. Цитоплазма эритроцита состоит из воды (60 %) и сухого остатка (40 %), содержащего около 95 % гемоглобина и 5 % других веществ. Наличие гемоглобина обусловливает желтую окраску отдельных эритроцитов свежей крови, а совокупность эритроцитов — красный цвет крови. При окрашивании мазка крови азур П-эозином по Романовскому —Гимзе большинство эритроцитов приобретают оранжево-розовый цвет (оксифильны), что обусловлено высоким содержанием в них гемоглобина. Функции: 1. Дыхательная — перенос кислорода в ткани и углекислого газа от тканей в легкие. 2. Регуляторная и защитная функции — перенос на поверхности различных биологически активных, токсических веществ, защитных факторов: аминокислот, токсинов, антигенов, антител и др. На поверхности эритроцитов часто может происходить реакция антиген-антитело, поэтому они пассивно участвуют в защитных реакциях. Обязательной составной частью популяции эритроцитов являются их молодые формы (1—5 %), называемые ретикулоцитами, или полихроматофильными эритроцитами. В них сохраняются рибосомы и эндоплазматическая сеть, формирующие зернистые и сетчатые структуры, которые выявляются при специальной суправитальной окраске (рис). При обычной гематологической окраске азур II -эозином они в отличие от основной массы эритроцитов, окрашивающихся в оранжево-розовый цвет (оксифилия), проявляют полихроматофилию и окрашиваются в сероголубой цвет. Гемоглобин относится к числу важнейших дыхательных белков, принимающих участие в переносе кислорода от легких к тканям. Он является основным компонентом эритроцитов крови, в каждом из них содержится примерно 280 млн молекул гемоглобина. Выделяют четыре формы гемоглобина: 1) оксигемоглобин; 2) метгемоглобин; 3) карбоксигемоглобин; 4) миоглобин. 3. Кровяные пластинки (тромбоциты). Классификация по степени зрелости. Размеры, строение, функции. Тромбоциты (кровяные пластинки от греч. thrombos – сгусток и cytos – клетка) мелкие дисковидные двояковыпуклые безъядерные постклеточные структуры диаметром 2–4 мкм, циркулирующие в крови. Они представляют собой окруженные мембраной и лишенные ядра фрагменты цитоплазмы мегакариоцитов. Они образуются в красном костном мозге в результате фрагментации участков цитоплазмы мегакариоцитов (гигантских клеток костного мозга), поступают в кровь, где находятся в количестве 2–4•109 /л крови, из этого числа 15% обновляется ежедневно. Средняя продолжительность жизни составляет 9–10 дней. Функции тромбоцитов 1. Остановка кровотечения при повреждении стенки сосуда (первичный гемостаз) – основная функция тромбоцитов. 2. Обеспечение свертывания крови (гемокоагуляция) – вторичный гемостаз 3. Участие в реакции заживления ран (главным образом повреждения сосудистой стенки) и воспаления. 4. Обеспечение нормальной функции сосудов, в частности их эндотелиальной выстилки – ангиотрофическая. Различают 5 основных форм тромбоцитов: 1. Юные – 10% 2. Зрелые – 80–85% 3. Старые – 5–10% 4. Дегенеративные – до 2% 5. Гигантские формы. Молодые формы тромбоцитов крупнее старых. Тромбоцит окружен плазмолеммой и включает светлую прозрачную наружную часть, называемую гиаломером, и центральную окрашенную часть, содержащую азурофильные гранулы –грануломер. Плазмолемма покрыта снаружи толстым (50–200 нм) слоем гликокаликса. Она содержит многочисленные рецепторы, опосредующие действие веществ. Активирующих или ингибирующих функции тромбоцитов, их адгезию и агрегацию. Сама плазмолемма образует инвагинации с отходящими канальцами, также покрытыми гликокаликсом. Гиаломер содержит две системы трубочек (канальцев) и большую часть элементов цитоскелета. Цитоскелет тромбоцитов представлен микротрубочками, микрофиламентами и промежуточными филаментами.– Микротрубочки в количестве 4–15 располагаются по периферии цитоплазмы и формируют мощный пучок (краевое кольцо) служащий каркасом и способствующий поддержанию формы тромбоцитов.– Микрофиламенты образованы актином. 4. Зернистые лейкоциты (гранулоциты). Классификация, их содержание, размеры, форма, строение, основные функции. Лейкоциты. Лейкоциты, или белые кровяные клетки, в свежей крови бесцветны. Число их составляет в среднем 4-9 • 109 л. Лейкоциты в кровяном русле и лимфе способны к активным движениям, могут переходить через стенку сосудов в соединительную ткань органов, где они выполняют основные защитные функции. По морфологическим признакам и биологической роли лейкоциты подразделяют на две группы: зернистые лейкоциты, или гранулоциты, и незернистые лейкоциты, или агранулоциты. У зернистых лейкоцитов выявляются специфическая зернистость (эозинофильная, базофильная или нейтрофильная) и сегментированные ядра. В соответствии с окраской специфической зернистости различают нейтрофильные, эозинофильные и базофильные гранулоциты. Ядро обычно дольчатое (сегментированное), однако незрелые их формы имеют палочковидное ядро. К гранулоцитам относятся нейтрофильные, эозинофильные и базофильные лейкоциты. Они образуются в красном костном мозге, содержат специфическую зернистость в цитоплазме и сегментированные ядра. Нейтрофильные гранулоциты— самая многочисленная группа лейкоцитов, составляющая 2,0—5,5 • 109 л крови. Их диаметр в мазке крови 10—12 мкм, а в капле свежей крови 7—9 мкм. В популяции нейтрофилов крови могут находиться клетки различной степени зрелости — юные, палочкоядерные и сегментоядерные. В цитоплазме нейтрофилов видна зернистость. Эозинофильные грамулоциты. Количество эозинофилов в крови составляет 0,02— 0,3 • 109 л. Их диаметр в мазке крови 12—14 мкм, в капле свежей крови — 9—10 мкм. В цитоплазме расположены органеллы — аппарат Гольджи (около ядра), немногочисленные митохондрии, актиновые филаменты в кортексе цитоплазмы под плазмолеммой и гранулы. Среди гранул различают азурофильные (первичные) и эозинофильные (вторичные). Базофильные гранулоциты. Количество базофилов в крови составляет 0—0,06 • 109/л. Их диаметр в мазке крови равен 11 — 12 мкм, в капле свежей крови — около 9 мкм. В цитоплазме выявляются все виды органелл — эндоплазматическая сеть, рибосомы, аппарат Гольджи, митохондрии, актиновые фила-менты. 5. Незернистые лейкоциты (агранулоциты). Классификация, их содержание, размеры, форма, строение, основные функции. К этой группе лейкоцитов относятся лимфоциты и моноциты. В отличие от гранулоцитов они не содержат в цитоплазме специфической зернистости, а их ядра не сегментированы. Лимфоциты в крови взрослых людей составляют 20—35% от общего числа лейкоцитов. Различают крупные (от 10 до 18 мкм), средние (от 6,5 до 10 мкм) и малые (от 4,5 до 6,5 мкм) формы. В крови человека циркулируют преимущественно мелкие лимфоциты, размер которых не превышает в диаметре 7-10 мкм. Большие лимфоциты встречаются в крови новорожденных и детей, у взрослых они отсутствуют. Большую часть всех лимфоцитов крови человека составляют малые лимфоциты. Для всех видов лимфоцитов характерно наличие интенсивно окрашенного ядра округлой или бобовидной формы. В цитоплазме лимфоцитов содержится небольшое количество азурофильных гранул (лизосом). Основной функцией лимфоцитов является участие в иммунных реакциях. Однако популяция лимфоцитов гетерогенна по характеристике поверхностных рецепторов и роли в реакциях иммунитета. Среди лимфоцитов различают три основных функциональных класса: B-лимфоциты, T-лимфоциты и т.н. нулевые лимфоциты. Моноциты. Эти клетки крупнее других лейкоцитов. Это самые большие белые клетки, их размер в диаметре – от 12 до 20 мкм. Красновато-фиолетовое ядро может иметь разные формы: бобовидную, в виде подковы, бабочки, гриба и др. Оно занимает равную с цитоплазмой или большую часть клетки. В крови человека количество моноцитов от 6 до 8 % от общего числа лейкоцитов. Ядра моноцитов встречаются бобовидные, подковообразные, редко — дольчатые. Цитоплазма моноцитов менее базофильна, чем цитоплазма лимфоцитов. Она имеет бледно-голубой цвет, но по периферии окрашивается несколько темнее, чем около ядра. В цитоплазме содержится различное количество очень мелких азурофильных зерен (лизосом), расположенных чаще около ядра. Характерно наличие пальцеобразных выростов цитоплазмы и образование фагоцитарных вакуолей. В цитоплазме расположено множество пиноцитозных везикул. Моноциты относятся к макрофагической системе организма, или к так называемой мононуклеарной фагоцитарной системе. 6. Иммунокомпетентные клетки. Т-лимфоциты: классификации, субпопуляции, участие в иммунных реакциях, антигеннезависимая и антигензависимая бласттрансформация, пролиферация и дифференцировка. Иммунокомпетентные клетки - клетки, способные специфически распознавать антиген и отвечать на него иммунной реакцией. Такими клетками являются Т- и В-лимфоциты, которые под влиянием чужеродных агентов дифференцируются в сенсибилизированный лимфоцит и плазматическую клетку. Т-лимфоциты – это сложная по составу группа клеток, которая происходит от полипотентной стволовой клетки костного мозга, а созревает и дифференцируется в тимусе. Т-лимфоциты разделяются на две субпопуляции: иммунорегуляторы и эффекторы. Задачу регуляции иммунного ответа выполняют Т-хелперы. Эффекторную функцию осуществляют Т-киллеры и естественные киллеры. В организме Т-лимфоциты обеспечивают клеточные формы иммунного ответа, определяют силу и продолжительность иммунной реакции. Т-система иммунитета включает тимус как центральный орган системы, различные субпопуляции Т-клеток , группу цитокинов , продуцируемых Т-клетками. Происхождение Т-лимфоцитов. Данный росток дифференцировки на территории костного мозга проходит только самый начальный этап развития: формирование предшественника Т-клеток (пре-Т-клеток) от лимфоидной стволовой клетки; основные события по созреванию различных субпопуляций клоноспецифических Т-клеток разворачиваются в тимусе. Стволовая гемопоэтическая клетка, мигрирующая в тимус, превращается (дифференцируется) под влиянием тимического микроокружения в Т-лимфоцит. Локализация. Обычно локализуются в так называемых Т- зависимых зонах периферических лимфоидных органов (периартикулярно в белой пульпе селезенки и паракортикальных зонах лимфоузлов). Функции. Т- лимфоциты распознают процессированный и представленный на поверхности антигенпредставляющих ( А ) клеток антиген. Они отвечают за клеточный иммунитет, иммунные реакции клеточного типа. Отдельные субпопуляции помогают В- лимфоцитам реагировать на Т- зависимые антигены выработкой антител. Тлимфоциты распознают антигены с помощью двух типов мембранных гликопротеинов- Т- клеточных рецепторов (семейство Ig- подобных молекул) и CD3, нековалентно связанных между собой. Их рецепторы, в отличие от антител и рецепторов В- лимфоцитов, не распознают свободно циркулирующие антигены. Выделяют три основные группы Т- лимфоцитов- помощники (активаторы), эффекторы, регуляторы. Антигеннезависимая дифференцировка протекает независимо от присутствия антигенов, в центральных органах иммунных реакций. Для В-лимфоцитов это красный костный мозг, для Т-лимфоцитов – тимус (поэтому Тлимфоциты на этой стадии развития называют еще тимоцитами) Унипотентные предшественники (пре-В-клетки или пре-Т-клетки) проходят здесь стадии: Лимфобласт (соответствует большому лимфоциту) незрелый-лимфоцит (средний) зрелый лимфоцит (малый). Главные события антигензависимой дифференцировки – приобретение лимфоцитами специфических поверхностных рецепторов. 7. Иммунокомпетентные клетки. В-лимфоциты: классификации, субпопуляции,рецепторы к антигенам, антигеннезависимая и антигензависимая бласттрансформация, пролиферация и дифференцировка. Иммунокомпетентные клетки - клетки, способные специфически распознавать антиген и отвечать на него иммунной реакцией. Такими клетками являются Т- и В-лимфоциты, которые под влиянием чужеродных агентов дифференцируются в сенсибилизированный лимфоцит и плазматическую клетку. Клетками гуморального иммунитета являются В-лимфоциты: 10-30% от всех лимфоцитов. Короткоживущие (от 2 суток до 6 месяцев). Развитие: у птиц проходят антигеннезависимую дифференцировку в фабрициевой сумке. У человека таким органом является красный костный мозг, где формируется группа малодифференцированных Влимфоцитов (кластер CD 19). В-лимфоциты являются основными клетками, участвующими в гуморальном иммунитете. У человека они образуются из СКК красного костного мозга, затем поступают в кровь и далее заселяют Взоны периферических лимфоидных органов — селезенки, лимфатических узлов, лимфоидные фолликулы многих внутренних органов. При действии антигена В-лимфоциты в периферических лимфоидных органах активизируются, пролиферируют, дифференцируются в плазмоциты, активно синтезирующие антитела различных классов, которые поступают в кровь, лимфу и тканевую жидкость. Дифференцировочные антигены Т-клеток. В процессе дифференцировки лимфоцитов на их поверхности появляются специфические мембранные молекулы гликопротеидов. Такие молекулы (антигены) можно обнаружить с помощью специфических моноклональных антител. Получены моноклональные антитела, которые реагируют лишь с одним антигеном клеточной мембраны. С помощью набора моноклональных антител можно идентифицировать субпопуляции лимфоцитов. Имеются наборы антител к дифференцировочным антигенам лимфоцитов человека. 8. Макрофаги: свободные и фиксированные. Участие в иммунных реакциях, понятие о монокинах (медиаторах), кооперации иммунокомпетентных клеток. Макрофаги- это гетерогенная специализированная клеточная популяция защитной системы организма. Различают две группы макрофагов- свободные и фиксированные. К свободным макрофагам относятся макрофаги рыхлой соединительной ткани, или гистиоциты; макрофаги серозных полостей; макрофаги воспалительных экссудатов; альвеолярные макрофаги легких. Макрофаги способны перемещаться в организме. Группу фиксированных макрофагов составляют макрофаги костного мозга и костной ткани, селезенки, лимфатических узлов, внутриэпидермальные макрофаги, макрофаги ворсин плаценты, ЦНС. Кооперация иммунокомпетентных клеток. Иммунная реакция организма может иметь различный характер, но всегда начинается с захвата антигена макрофагами крови и тканей или же со связывания со стромой лимфоидных органов. Нередко антиген адсорбируется также на клетках паренхиматозных органов. В макрофагах он может полностью разрушаться, но чаше подвергается лишь частичной деградации. В частности, большинство антигенов в лизосомах фагоцитов в печение часа подвергается ограниченной денатурации и протеолизу. Оставшиеся от них пептиды (как правило, два-три остатка аминокислот) комплексируются с экспрессированными на внешней мембране макрофагов молекулами МНС. Монокины - клеточные медиаторы воспаления, которые образуются моноцитами/макрофагами на фоне антигенной стимуляции, а некоторые монокины могут продуцироваться другими клетками - лимфоцитами, гепатоцитами, глиальными клетками и др. 9. Лимфа. Лимфоплазма и форменные элементы. Связь с кровью, понятие о рециркуляции лимфоцитов. Лимфа состоит из лимфоплазмы и форменных элементов, в основном лимфоцитов (98 %), а также моноцитов, нейтрофилов, иногда эритроцитов. Лимфоплазма образуется посредством проникновения (дренажа) тканевой жидкости в лимфатические капилляры, а затем отводится по лимфатическим сосудам различного калибра и вливается в венозную систему. По пути движения лимфа проходит через лимфатические узлы, в которых она очищается от экзогенных и эндогенных частиц, а также обогащается лимфоцитами. По качественному составу лимфа подразделяется на: периферическую лимфу — до лимфатических узлов; промежуточную лимфу — после лимфатических узлов; центральную лимфу — лимфа грудного протока. В области лимфатических узлов происходит не только образование лимфоцитов, но и миграция лимфоцитов из крови в лимфу, а затем с током лимфы они снова попадают в крови и так далее. Такие лимфоциты составляют рециркулирующий пул лимфоцитов. Функции лимфы: дренирование тканей; обогащение лимфоцитами; очищение лимфы от экзогенных и эндогенных веществ. Лимфоциты постоянно рециркулируют из крови в ткани, и обратно. Рециркуляцией называют процесс перехода лимфоцитов из кровяного русла в органы, оттуда в лимфу и вновь в кровоток. При этом клетки, покинувшие конкретный лимфатический узел, возвращаются в любой узел или селезенку, что и обеспечивает перемешивание рециркулирующих лимфоцитов. Лимфоциты переходят из кровотока в лимфоидные образования обычные посткапиллярные венулы, однако, этот переход обычно осуществляется преимущественно через специализированные участки венозного русла – венулы с высоким эндотелием. В лимфоузлах эти сосуды находятся главным образом в паракортикальной области и иногда в корковой. Вместе с тем часть лимфоцитов, а в первую очередь Т-клетки, поступают в региональный лимфатический узел из дренируемой им области по приносящим лимфатическим сосудам. 10. Унитарная теория кроветворения А. А. Максимова и ее современная трактовка. Унитарная теория кроветворения - все клетки крови развиваются из единой одной и той же родоначальной клетки (морфологически это малый лимфоцит). Основоположником современной унитарной теории кроветворения является отечественный гистолог Максимов (работал на кафедре гистологии ВМА в С-Петербурге). Еще в 1907 году Максимов утверждал, что все клетки крови развиваются из единой одной и той же родоначальной клетки; мало того, он назвал эту клетку — морфологически это малый лимфоцит. Однако имеющиеся в то время методы исследований не позволяли экспериментально доказать верность этой теории. Максимов в ходе гемоцитопоэза клетки крови подразделял на 4 группы: 1 группа — клетки с неограничанной возможностью превращений, т.е. родоначальная клетка, способная развиваться и превратиться в любой форменный элемент крови. 2 группа — клетки с частично ограниченный способностью развиваться в ту или иную форму клеток крови. 3 группа — клетки со строго ограничанной возможностью развития. 4 группа — клетки крови не способные изменяться. Последующие исследования показали верность унитарной теории кроветворения Максимова. В настоящее время общепринятой является унитарная теория кроветворения, на основании которой разработана схема кроветворения (И. Л. Чертков и А. И. Воробьев, 1973 г.). В процессе поэтапной дифференцировки стволовых клеток в зрелые форменные элементы крови в каждом ряду кроветворения образуются промежуточные типы клеток, которые в схеме кроветворения составляют классы клеток. Всего в схеме кроветворения различают 6 классов клеток: 1 класс - стволовые клетки; 2 класс - полустволовые клетки; 3 класс - унипотентные клетки; 4 класс - бластные клетки; 5 класс - созревающие клетки; 6 класс - зрелые форменные элементы. Морфологическая и функциональная характеристика клеток различных классов схемы кроветворения. 11. Этапы кроветворения в эмбриональный и постэмбриональный периоды развития и их биологической значение. Гемопоэз – развитие крови. Различают эмбриональный гемопоэз, который происходит в эмбриональный период и приводит к развитию крови как ткани, и постэмбриональный гемопоэз, который представляет собой процесс физиологической регенерации крови. Эмбриональный гемопоэз. В развитии крови как ткани в эмбриональный период можно выделить 3 основных этапа: Мезобластический (желточный), когда начинается развитие клеток крови во внезародышевых органах и появляется первая регенерация стволовых клеток крови. (с 3-й по 9-ю неделю). Печеночный (гепатотимусолиенальный), который начинается в печени с 5-6-й недели развития плода, когда печень становится основным органом гемопоэза, в ней образуется вторая регенерация СКК. Кроветворение в печени достигает максимума через 5 мес и завершается перед рождением. Медуллярный (костномозговой) (медулло-тимусолимфоидный)– появление третьей генерации СКК в костном мозге, где гемопоэз начинается с 10-й недели и постепенно нарастает к рождению, а после рождения красный костный мозг становится центральным органом гемопоэза. Постэмбриональный гемопоэз. Постэмбриональный гемопоэз представляет собой процесс физиологической регенерации крови (клеточное обновление), который компенсирует физиологическое разрушение дифференцированных клеток. Постэмбриональное кроветворение совершается в миелоидной ткани костного мозга и лимфоидной ткани тимуса, селезенки и лимфатических узлов и является физиологической регенерацией крови, т. е. компенсирует естественную убыль клеток в результате их старения и разрушения. В миелоидной ткани образуются эритроциты, гранулоциты, моноциты, кровяные пластинки, в лимфоидной ткани —лимфоциты. 12. Понятие о стволовых и полустволовых клетках, дифферонах. Взаимоотношения стромальных и кроветворных элементов. Стволовые клетки. В красном костном мозге находятся так называемые стволовые клетки — предшественницы всех форменных элементов крови, которые (в норме) поступают из костного мозга в кровяное русло уже полностью зрелыми. Полустволовые клетки (ПСК) - клетки предшественники миелопоэза, клетки предшественники лимфопоэза. Дифферон (гистогенетический ряд) - совокупность клеточных форм, составляющих ту или иную линию дифференцировки. В диффероне различают: стволовые клетки - клетки-предшественницы - зрелые клетки, достигшие состояния окончательной (терминальной) дифференцировки. Стволовые клетки самоподдерживающаяся популяция клеток, способных дифференцироваться в нескольких направлениях и формировать различные клеточные типы. Так, стволовые эпендимные клетки ЦНС дают начало разным нейронам и глиоцитам. Стволовые клетки обладают высокими пролиферативными потенциями, но, как правило, делятся редко. Клеткипредшественницы. По мере дифференцировки пролиферативные потенции клеток постепенно уменьшаются. Выделяют наиболее раннюю стадию клеток-предшественниц -коммитированные, или полустволовые клетки. Зрелые клетки. Ими заканчивается гистогенетический ряд. Стволовые клетки. Стволовыми клетками эмбриона, плода и взрослого организма считаются клетки, которые при определённых условиях способны длительное время воспроизводить себе подобные клетки и в течение жизни давать начало специализированным клеткам, образующих ткани и органы тела. Выделяют тотипотентные и плюрипотентные стволовые клетки. Стромальные ретикулярные и гемопоэтические элементы. Для миелоидной и всех разновидностей лимфоидной ткани характерно наличие стромальных ретикулярных и гемопоэтических элементов, образующих единое функциональное целое. В тимусе имеется сложная строма, представленная как соединительнотканными, так и ретикулоэпителиальными клетками. Эпителиальные клетки секретируют особые вещества — тимозины, оказывающие влияние на дифференцировку из СКК Т-лимфоцитов. В лимфатических узлах и селезенке специализированные ретикулярные клетки создают микроокружение, необходимое для пролиферации и дифференциров-ки в специальных Т- и В-зонах Т- и В-лимфоцитов и плазмоцитов. СКК являются полипотентными предшественниками всех клеток крови 13. Эритроцитопоэз, тромбоцитопоэз. В эритроцитопоэзе самой молодой клеткой является эритробласт (ее называют также проэритробластом), который имеет бластную структуру и обычно круглое ядро. Цитоплазма при окраске темно-синяя, располагается узким ободком, часто образует своеобразные выросты. В отношении клеток эритрокариоцитарного ряда нет единой номенклатуры. Их называют как нормобластами, так и эритробластами. Поскольку для других рядов термин «бласт» применяется лишь для клеток-родоначальниц того или иного ростка (отсюда и название «бласт» – росток), все клетки, являющиеся потомством эритробласта, должны иметь в названии окончание «цит». Поэтому термин «нормобласты» был заменен на «нормоциты». За эритробластом появляется пронормоцит, который отличается от эритробласта более грубым строением ядра, хотя оно и сохраняет правильную структуру хроматиновых нитей. Диаметр ядра меньше, чем у эритробласта, ободок цитоплазмы шире, видна перинуклеарная зона просветления. При изучении миелограммы пронормоцит легко спутать с эритробластом. В связи с трудностью разделения этих клеток некоторые авторы предлагают в практической гематологии их вообще не дифференцировать. Далее следует базофильный нормоцит, у которого грубоглыбчатое ядро имеет колосовидную структуру, а цитоплазма окрашена в темно-синий цвет. Следующий – полихроматофильный нормоцит отличается еще более плотной структурой ядра; цитоплазма занимает большую часть клетки и имеет базофильную (за счет структур, содержащих РНК) и оксифильную (в связи с появлением достаточного количества гемоглобина) окраску. Тромбоцитопоэз. Самой молодой клеткой тромбоцитопоэза является мегакариобласт – одноядерная небольшая клетка с крупным бластным ядром, хроматинные нити которого толще и грубее, чем у эритробласта; в ядре могут быть видны 1-2 темно-синие нуклеолы. Цитоплазма беззернистая, темно-синего цвета, отростчатая, узким ободком окружает ядро. Промегакариоцит возникает в результате нескольких эндомитозов. Ядро полиморфное, с грубым строением хроматина; цитоплазма темно-синяя, беззернистая. Зрелый мегакариоцит отличается от промегакариоцита большим ядром. Цитоплазма имеет сине-розовую окраску, содержит азурофильную красноватую зернистость. Внутри мегакариоцита формируются тромбоциты. В мазке крови можно видеть и распадающиеся мегакариоциты, окруженные кучками тромбоцитов. При тромбоцитолитических состояниях отшнуровка тромбоцитов может происходить и на стадии промегакариоцита; тромбоциты при этом лишены азурофильной субстанции, но активно участвуют в гемостазе. 14. Лейкоцитопоэз (гранулоцитопоэз). Лейкоцитопоэз включает гранулоцитопоэз, лимфоцитопоэз и моноцитопоэз. В гранулоцитарном ряду миелобласт представляет первую морфологически различимую клетку. Он имеет нежноструктурное ядро, единичные нуклеолы. Форма ядра круглая, размеры чуть меньше, чем у эритробласта. Миелобласт отличается от недифференцируемых бластов из класса клеток-предшественниц наличием зернистости в цитоплазме; форма клетки чаще круглая, ровная. На следующей стадии гранулоцитопоэза обнаруживается промиелоцит – нейтрофильный, эозинофильный и базофильный. Круглое или бобовидное ядро промиелоцита больше ядра миелобласта почти вдвое, хотя эта клетка и не является полиплоидией; оно часто располагается эксцентрично, и в нем можно видеть остатки нуклеол. Структура хроматина уже утрачивает нежное нитчатое строение бластных клеток, хотя и не приобретает грубоглыбчатого строения. Площадь цитоплазмы примерно равна площади ядра; цитоплазма обильно насыщена зернистостью, имеющей характерные для каждого ряда особенности. Для нейтрофильного ряда промиелоцит является самой зернистой клеткой. Его зернистость полиморфная: крупная и мелкая; окрашивается и кислыми, и основными красителями. В промиелоците зернистость часто располагается также на ядре. Зернистость эозинофильного промиелоцита, обладая характерной для эозинофилов однотипностью зерен (типа кетовой икры), вместе с тем окрашивается как кислыми, так и основными красителями. Базофильный промиелоцит имеет крупную полиморфную базофильную зернистость. 15. Лейкоцитопоэз (лимфоцитопоэз и моноцитопоэз). Лимфоцитопоэз. Из полипотентной гемопоэтической стволовой клетки происходит общая клетка-предшественница лимфоцитопоэза (КПЛ). Как и другие предшественницы, эта клетка не имеет специфических морфологических характеристик, хотя обладает большой способностью к преобразованиям. После деления класс лимфоидных стволовых клеток (СК-Л), определяющих образование популяции Т-лимфоцитов, покидает костный мозг и мигрирует в кору тимуса, где имеется очень специфичная микросреда, в которой СК-Л превращаются в Т-лимфоциты. Из В-лимфоцитов возникают, пройдя стадию плазмобласта, плазматические клетки (ПК) и В-клетки памяти; из Тлимфоцитов происходят следующие виды лимфоцитов: супрессоры, хелперы, киллеры и Т-клетки памяти. Большинство В-лимфоцитов циркулирует в крови и заканчивает свою дифференцировку и созревание в селезенке, лимфатических узлах и аналогах сумки Фабриция. Моноцитопоэз. Он начинается с образования промонобласта (Пмб) из общей стволовой клетки для гранулоцитопоэза, эритроцитопоэза, моноцитопоэза и мегакариоцитопоэза (КОЕ-С). Промонобласт — это большая, слегка базофильная клетка с крупным круглым ядром и плохо развитыми органеллами. После деления из промонобластов образуются монобласты (Мб) — клетки с почкообразным ядром и несколькими азурофильными гранулами. Из монобластов после деления образуются моноциты (Мц), которые выходят в кровеносное русло. VI. ТКАНИ ВНУТРЕННЕЙ СРЕДЫ. 1. Морфофункциональная характеристика и классификация соединительных тканей. Источники развития. Гистогенез. Вклад отечественных и зарубежных учёных в изучение соединительных тканей (А. А. Максимов, А. А. Заварзин, А. В. Румянцев, Г. К. Хрущёв, В. Г. Елисеев). Соединительные ткани — это комплекс мезенхимных производных, состоящий из клеточных дифферонов и большого количества межклеточного вещества (волокнистых структур и аморфного вещества), участвующих в поддержании гомеостаза внутренней среды и отличающихся от других тканей меньшей потребностью в аэробных окислительных процессах. Соединительная ткань участвует в формировании стромы органов, прослоек между другими тканями, дермы кожи, скелета. Классификация соединительных тканей. Разновидности соединительной ткани различаются между собой составом и соотношением клеток, волокон, а также физико-химическими свойствами аморфного межклеточного вещества. Соединительные ткани подразделяются на собственно соединительную ткань (волокнистые соединительные ткани и соединительные ткани со специальными свойствами) и скелетные ткани. Последние в свою очередь подразделяются на три разновидности хрящевой ткани (гиалиновая, эластическая, волокнистая), две разновидности костной ткани (фиброзно-волокнистая и пластинчатая), а также цемент и дентин зуба. Общие принципы организации соединительных тканей. Главными компонентами соединительных тканей являются производные клеток — волокнистые структуры коллагенового и эластического типов, основное (аморфное) вещество, играющее роль интегративно-буферной метаболической среды, и клеточные элементы, создающие и поддерживающие количественное и качественное соотношение состава неклеточных компонентов. Гистогенез соединительных тканей. Различают эмбриональный и постэмбриональный гистогенез соединительных тканей. В процессе эмбрионального гистогенеза мезенхима приобретает черты тканевого строения раньше закладки других тканей. Этот процесс в различных органах и системах происходит неодинаково и зависит от их неодинаковой физиологической значимости на различных этапах эмбриогенеза. Постэмбриональный гистогенез в нормальных физиологических условиях происходит медленнее и направлен на поддержание тканевого гомеостаза, пролиферацию малодифференцированных клеток и замену ими отмирающих клеток. Илья Ильич Мечников (1845-1916 гг.) еще при жизни был признан Гордостью России. Врач, многогранный ученый, обнаружил и описал явление фагоцитоза и сформулировал основные положения патологии воспаления, создав фагоцитарную теорию иммунитета, за что в 1908 г. вместе с П. Эрлихом был награжден Нобелевской премией в области физиологии и медицины. Александр Александрович Максимов (1874- 1928 гг.) — выдающийся российский учёный, гистолог и эмбриолог, основатель экспериментальной морфологии соединительной ткани и крови. Он впервые высказал идею о существовании стволовой клетки, единой для кроветворных, а возможно, и для других элементов тканей внутренней среды в постнатальном (после рождения) онтогенезе. В изучение функций и структуры соединительной ткани большой вклад внёс Алексей Алексеевич Заварзин. В своих сочинениях (Очерки эволюционной гистологии крови и соединительной ткани (1945-1947г.г.), Руководство по гистологии (в соавторстве с С. И. Щелкуновым)– 1954 г.) он справедливо считал, что рассматривать гистогенез соединительной ткани необходимо в неразрывном единстве с гистогенезом крови. В 1975 г. вышла в свет монография Н. Г. Хрущова «Гистогенез соединительной ткани», не большая по объему, но очень ёмкая по глубине изложенной в ней мысли о том, что в постнатальном онтогенезе млекопитающих существуют две популяции фибробластов, имеющих разное биологическое предназначение. 2. Морфофункциональная характеристика клеток рыхлой волокнистой соединительной ткани. Рыхлая волокнистая соединительная ткань обнаруживается во всех органах, так как она сопровождает крсвеносные и лимфатические сосуды и образует строму многих органов. Не смотря на наличие органных особенностей, строение рыхлой волокнистой соединительной ткани в различных органах имеет сходство. Она состоит из клеток и межклеточного вещества. Различают следующие клетки рыхлой волокнистой соединительной ткани: 1. Фибробласты – наиболее многочисленная группа клеток, различных по степени дифференцировки, характеризующаяся прежде всего способностью синтезировать фибриллярные белки (коллаген, эластин) и гликозаминогликаны с последующим выделением их в межклеточное вещество. 2. Макрофаги – блуждающие, активно фагоцитирующие клетки. Цитолемма макрофагов образует глубокие складки и длинные микровыросты, с помощью которых эти клетки захватывают инородные частицы. Как правило, имеют одно ядро. Цитоплазма базофильна, богата лизосомами, фагосомами и пиноцитозными пузырьками, содержит умеренное количество митохондрий, гранулярной эндоплазматической сети, комплекса Гольджи, включений гликогена, липидов и др. 3. Плазматические клетки (плазмоциты). Их величина колеблется от 7 до 10 мкм. Форма клеток округлая или овальная. Ядра относительно небольшие, круглой или овальной формы, расположены эксцентрично. Цитоплазма резко базофильна, содержит хорошо развитую гранулярную эндоплазматическую сеть, в которой синтезируются белки (антитела). 4. Тканевые базофилы (тучные клетки). Клетки их имеют разнообразную форму, иногда с короткими широкими отростками, что обусловлено способностью их к амебоидным движениям. В цитоплазме находится специфическая зернистость (синего цвета), напоминающая гранулы базофильных лейкоцитов. В ней содержится гепарин, гиалуроновая кислота, гистамин и серотонин. Органеллы тучных клеток развиты слабо. 5. Адипоциты (жировые клетки) – располагаются группами, реже – поодиночке. Накапливаясь в больших количествах, эти клетки образуют жировую ткань. 6. Пигментные клетки – имеют короткие, непостоянной формы отростки. Эти клетки содержат в своей цитоплазме пигмент меланин, способный поглощать УФЛ. 7. Адвентициальные клетки — малоспециализированные клетки, сопровождающие кровеносные сосуды. Они имеют уплощенную или веретенообразную форму со слабобазофильной цитоплазмой, овальным ядром и слаборазвитыми органеллами. 8. Перициты имеют отросчатую форму и в виде корзинки окружают кровеносные капилляры, располагаясь в расщелинах их базальной мембраны. 9. Лейкоциты мигрируют в соединительную ткань из крови. Межклеточное вещество состоит из основного вещества и расположенных в них волокон – коллагеновых, эластических и ретикулярных. Коллагеновые волокна в рыхлой неоформленной волокнистой соединительной ткани располагаются в различных направлениях в виде скрученных округлых или уплощенных тяжей толщиной 1-3 мкм и более. Внутренняя структура коллагенового волокна определяется фибриллярным белком – коллагеном, который синтезируется в рибосомах гранулярной эндоплазматической сети фибробластов. Эластические волокна – их форма округлая или уплощенная, широко анастомозируют друг с другом. Толщина эластических волокон обычно меньше коллагеновых. Основным химическим компонентом эластических волокон является глобулярный белок эластин, синтезируемый фибробластами. Ретикулярные волокна относятся к типу коллагеновых волокон, но отличаются меньшей толщиной, ветвистостью и анастомозами. Содержат повышенное количество углеводов, которые синтезируются ретикулярными клетками и липидов. Основное вещество – это студнеобразная гидрофильная среда, в образовании которой важную роль играют фибробласты. В его состав входят сульфатированные (хондроитинсерная кислота, кератин-сульфат, и др.) и несульфатированные (гиалуроновая кислота) гликозаминогликаны, которые обусловливают консистенцию и функциональные особенности основного вещества. Кроме указанных компонентов, в состав основного вещества входят липиды, альбумины и глобулины крови, минеральные вещества (соли натрия, калия, кальция и др.). 3. Межклеточное вещество соединительной ткани. Общая характеристика и строение. Фибробласты и их роль в образовании межклеточного вещества. Межклеточное вещество состоит из основного вещества и расположенных в них волокон – коллагеновых, эластических и ретикулярных. Коллагеновые волокна в рыхлой неоформленной волокнистой соединительной ткани располагаются в различных направлениях в виде скрученных округлых или уплощенных тяжей толщиной 1-3 мкм и более. Внутренняя структура коллагенового волокна определяется фибриллярным белком – коллагеном, который синтезируется в рибосомах гранулярной эндоплазматической сети фибробластов. Эластические волокна – их форма округлая или уплощенная, широко анастомозируют друг с другом. Толщина эластических волокон обычно меньше коллагеновых. Основным химическим компонентом эластических волокон является глобулярный белок эластин, синтезируемый фибробластами. Ретикулярные волокна относятся к типу коллагеновых волокон, но отличаются меньшей толщиной, ветвистостью и анастомозами. Содержат повышенное количество углеводов, которые синтезируются ретикулярными клетками и липидов. Фибробласты – наиболее многочисленная группа клеток, различных по степени дифференцировки, характеризующаяся прежде всего способностью синтезировать фибриллярные белки (коллаген, эластин) и гликозаминогликаны с последующим выделением их в межклеточное вещество. 4. Плотные волокнистые соединительные ткани, разновидности, строение и функции. Строение сухожилий и связок. Плотные волокнистые соединительные ткани характеризуются относительно большим количеством плотно расположенных волокон и незначительным количеством клеточных элементов и основного аморфного вещества между ними. В зависимости от характера расположения волокнистых структур эта ткань подразделяется на плотную неоформленную и плотную оформленную соединительную ткань. Плотная неоформленная соединительная ткань характеризуется неупорядоченным расположением волокон. Она образует капсулы, надхрящницу, надкостницу, сетчатый слой дермы кожи. Плотная оформленная соединительная ткань содержит строго упорядоченно расположенные волокна, толщина которых соответствует тем механическим нагрузкам, в которых функционирует орган. Оформленная соединительная ткань встречается, например, в сухожилиях, которые состоят из толстых, параллельно расположенных пучков коллагеновых волокон. При этом каждый пучок, отграниченный от соседнего слоем фиброцитов, называется пучком Iго порядка. Несколько пучков I порядка, разделенные прослойками рыхлой волокнистой соединительной ткани, называются пучком II-го порядка. Прослойки рыхлой волокнистой соединительной ткани называются эндотенонием. Пучки II порядка объединяются в более толстые пучки III-го порядка, окруженные более толстыми прослойками рыхлой волокнистой соединительной ткани, называемыми перитенонием. Пучки III порядка могут являться сухожилием, а в более крупных сухожилиях могут объединяться в пучки IV-го порядка, которые также окружены перитенонием. Эндотеноний и перитеноний содержат питающие сухожилие кровеносные сосуды, нервы и проприоцептивные нервные окончания. Сухожилие – удлинённые образования, которые связывают поперечнополосатую соматическую мышцу с костью. Сухожилия являются органами, поскольку состоят из нескольких видов тканей, в частности ПВОСТ и РВСТ. а) Пучки волокон первого порядка состоят из тесно прилегающих друг к другу коллагеновых волокон. Между этими пучками располагаются фибробласты и фиброциты (с преобладанием фиброцитов). б) Пучки первого порядка объединяются в пучки второго порядка, разделенные прослойками РВСТ - эндотенонием. в) Пучки второго порядка образуют пучки третьего порядка, покрытые перитенонием. Иногда пучком третьего порядка является само сухожилие. Связки соединяют кости друг с другом и по строению сходны с сухожилиями, отличаясь от них несколько менее строго ориентированным расположение коллагеновых волокон. В большинстве связок преобладают коллагеновые волокна. 5. Макрофаги. Понятие о системе мононуклеарных фагоцитов Макрофаги- это гетерогенная специализированная клеточная популяция защитной системы организма. Различают две группы макрофагов-свободные и фиксированные.К свободным макрофагам относятся макрофаги рыхлой соединительной ткани, или гистиоциты; макрофаги серозных полостей; макрофаги воспалительных экссудатов; альвеолярные макрофаги легких. Макрофаги способны перемещаться в организме. Группу фиксированных макрофагов составляют макрофаги костного мозга и костной ткани, селезенки, лимфатических узлов, внутриэпидермальные макрофаги, макрофаги ворсин плаценты, ЦНС. Обычно макрофаги имеют одно ядро. Ядра макрофагов небольшого размера, округлые, бобовидные или неправильной формы. В них содержатся крупные глыбки хроматина. Цитоплазма базофильна, богата лизосомами, фагосомами и пиноцитозными пузырьками, содержит умеренное количество митохондрий, гранулярную эндоплазматическую сеть, аппарат Гольджи, включения гликогена, липидов и др.Формы проявления защитной функции макрофагов: 1) поглощение и дальнейшее расщепление или изоляция чужеродного материала; 2) обезвреживание его при непосредственном контакте; 3) передача информации о чужеродном материале иммунокомпетентным клеткам, способным его нейтрализовать; 4) оказание стимулирующего воздействия на другую клеточную популяцию защитной системы организма. Количество макрофагов и их активность особенно возрастают при воспалительных процессах. Макрофаги вырабатывают факторы, активирующие выработку иммуноглобулинов Влимфоцитами, Понятие о макрофагической системе.К этой системе относятся совокупность всех клеток, обладающих способностью захватывать из тканевой жидкости организма инородные частицы, погибающие клетки, неклеточные структуры, бактерии и др. Фагоцитированный материал подвергается внутри клетки ферментативному расщеплению, благодаря чему ликвидируются вредные для организма агенты, возникающие местно или проникающие извне. И.И. Мечников первым пришел к мысли о том, что фагоцитоз, возникающий в эволюции как форма внутриклеточного пищеварения и закрепившийся за многими клетками, одновременно является важным защитным механизмом. 6. Соединительные ткани со специальными свойствами (ретикулярная, жировая, пигментная, слизистая). Соединительные ткани – это комплекс мезенхимных производных, состоящий из клеточных дифферонов и большого количества межклеточного вещества (волокнистых структур и аморфного вещества). Общая характеристика соединительных тканей: - самая распространенная ткань (составляет более 50% массы тела человека); - развивается из мезенхимы; - состоит из клеток и межклеточного вещества; - ткань полидифферонная; - клетки аполярны; - в составе соединительной ткани есть кровеносные сосуды; - хорошо регенерируют. Общие функции соединительной ткани:1) трофическая;2) функция механической защиты (кости черепа);3) опорно-механическая (костная, хрящевая ткани, сухожилия, апоневрозы);4) формообразующая (склера глаза придает глазу определенную форму);5) защитная (фагоцитоз и иммунологическая защита);6) пластическая (способность адаптироваться к новым условиям внешней среды, участие в заживлении ран);7) участие в поддержании гомеостаза организма. Соединительные ткани со специальными свойствами относятся к группе собственно соединительных тканей. Среди соединительных тканей со специальными свойствами выделяют: жировую, ретикулярную и слизистую. Они характеризуются преобладанием однородных клеток. Жировая ткань: Это скопление жировых клеток, встречающихся во многих органах. Различают белую (широко распространена в организме) и желтую (встречается только детей до 1ого месяца жизни) жировые ткани. Белая жировая ткань располагается под кожей, Жировые клетки в дольках тесно прилежат друг к другу, а между ними располагаются фибробласты, лимфоидные элементы, тканевые базофилы и так же тонкие коллагеновые волокнаЖировая ткань играет роль депо жира и воды, механическую, трофические функции. Во время голодания жировая ткань быстро теряет запасы жира. И жировые клетки приобретают звездчатую форму, когда в норме у них округлая форма. Бурая (желтая) жировая ткань встречается на шее, около лопатокОна состоит из жировых клеток, густо оплетенных гемокапиллярами. Клетки содержат много митохондрий, которые содержат железосодержащие пигменты – цитохромы, которые придают желтый цвет Ретикулярная ткань: Имеет сетевидное строение и состоит из отростчатых ретикулярных клеток и ретикулярных волокон. Большинство ретикулярных клеток связываются в волокнами, образуя 3-ех мерную сеть. Ретикулярные волокна представляют собой продукт синтеза ретикулярных клеток. Среди них различают собственно ретикулярные и преколлагеновые волокна. Ретикулярные волокна содержат больше серы, липидов и углеводов по сравнению с коллагеновыми. Функция ретикулярной ткани: основа стромы органов кроветворения и создает микроокружение для развивающихся клеток крови. 3. Слизистая ткань: В норме встречается только у зародыша в пупочном канатике плода. Клеточные элементы представлены гетерогенной группой клеток, дифференцирующихся из мезенхимы. Они образуют дифферон: фибробласты – миофибробласты – гладкие миоциты. Они отличаются способностью синтезировать виментин, десмин, актин и миозин, коллаген 4 типа. В первой половине беременности между клетками содержится большое количество гиалуроновой кислоты. А на поздних этапах развития между клетками появляются рыхло расположенные коллагеновые волокна. 7. Хрящевые ткани. Общая характеристика. Виды хрящевых тканей. Строение клеток и межклеточного вещества. Возрастные изменения. Хрящеваяи костная тканиразвиваются из склеротомной мезенхимы, Хрящевые ткани. Классифицируются на гиалиновую, эластическую и волокнистую. В основу классификации положены особенности организации межклеточного веществаКлетки хрящевой ткани).Составляют дифферон хондроцитов: стволовая клетка, полустволовая клетка (прехондробласт), хондробласт, хондроцит. Хондроциты располагаются в хрящевых лакунах. В лакуне вначале находится 1 хондроцит, потом, в процессе его митотического деления, образуется 2, 4, 6 и т. д. клеток. Все они находятся в одной лакуне и образуют изогенную группу хондроцитов.Хондроциты изогенной группы делятся на 3 типа: I, II, III. Хондроциты I типаобладают способностью к митотическому делению, содержат комплекс Гольджи, митохондрии, гранулярную ЭПС и свободные рибосомы, имеют крупное ядро и небольшое количество цитоплазмы (большое ядерно-цитоплазматическое отношение). Эти хондроциты располагаются в молодом хряще. Хондроциты II типарасполагаются в зрелом хряще, ядерно-цитоплазматическое отношение их несколько уменыпается, так как увеличивается объем цитоплазмы; они утрачивают способность к митозуХондроциты III типанаходятся в старом хряще, утрачивают способность к синтезу гликозаминогликанов и вырабатывают только белки, поэтому межклеточное вещество вокруг них окрашивается оксифильно. Межклеточное вещество хрящевой ткани.Содержит органические вещества (преимущественно коллаген II типа), гликозаминогликаны, протеогликаны и белки неколлагено- вого типа. Чем больше протеогликанов, тем более гидрофильно межклеточное вещество, тем оно более упруго и более проницаемо. Через основное вещество со стороны надхрящницы диффузно проникают газы, молекулы воды, ионы солей и микромолекулы. Однако макромолекулы не проникаютВ основном веществе хряща имеются коллагеновые волокна, состоящие из коллагена II типаВ межклеточном веществе хрящевой ткани отсутствуют кровеносные и лимфатические сосуды, поэтому питание хрящевой ткани осуществляется путем диффузного поступления веществ со стороны сосудов надхрящницыВозрастные изменения хрящевой ткани.Наибольшие изменения наблюдаются в пожилом возрасте, когда уменьшается количество хондробластов в надхрящнице и число делящихся хрящевых клеток. В хондроцитах уменьшается количество гранулярной ЭПС, комплекса Гольджи и митохондрий, утрачивается способность хондроцитов к синтезу гликозаминогликанов и протеогликанов. Снижение количества протеогликанов приводит к уменьшению гидрофильности хрящевой ткани, ослаблению проницаемости хряща и поступлению питательных веществ. Это приводит к обызвествлению хряща, проникновению в него кровеносных сосудов и образованию костного вещества внутри хрящевого. 8. Строение сустава. Строение суставного хряща. Суставы соединяют кости между собой и определяют характер и степень движений между ними. Они расположены по краям костей, покрыты и окружены соединительными тканями. Суставы можно разделить на диартрозы, в которых происходят свободные движения костей, и синартрозы (греч. syn — вместе + arthrosis — сочленение), в которых движения ограничены или отсутствуют. В синостозе кости связаны костной тканью, и движения отсутствуют. оллагеновые волокна суставной поверхности хряща располагаются в виде готических арок, которые служат конструкциями для оптимального распределения сил, создаваемых давлением на эту ткань. Суставные капсулы (сумки) диартрозов варьируют по структуре в соответствии с особенностями сустава. Обычно, однако, такая капсула включает два слоя — наружный фиброзный слой и внутренний синовиальный слой. Синовиальный слой образован двумя типами клеток. Один напоминает фибробласты, а другой по строению и поведению сходен с макрофагами. Фиброзный слой образован плотной соединительной тканью. Клетки (хондроциты) составляют не более 10% массы хряща. Основной объем в хрящевой ткани приходится на межклеточное вещество. Аморфное вещество достаточно гидрофильно, что позволяет доставлять клеткам питательные вещества путем диффузии из капилляров надхрящницы. 9. Морфофункциональная характеристика и классификация костных тканей. Клетки и межклеточное вещество. Возрастные изменения. Костная ткань - специализированный вид соединительной ткани с очень высокой степенью минерализации межклеточного вещества. Костная ткань .развивается из мезенхимы двумя способами: непосредственно из мезенхимы (прямой остеогенез) либо на месте сформированного ранее гиалинового хряща (непрямой остеогенез). Костная ткань, как и все разновидности соединительной, состоит из клеток и межклеточного вещества. Клетки кости - остеобласты, остеоциты и остеокласты. В составе межклеточного вещества - оссеиновые волокна (коллагеновые волокна 1-го типа) и основное вещество(содержит неколлагеновые белки, липиды, гликопротеиды, гликозаминогликаны и протеогликаны). Остеобласты - молодые клетки, создающие межклеточное вещество кости. В образующейся кости они покрывают почти всю поверхность костной балки. В сформированной кости остеобласты встречаются только в глубоких слоях надкостницы, Остеоциты - наиболее многочисленные клетки костной ткани. Они имеют отростчатую форму, округлое плотное ядро и слабобазофильную цитоплазму. Остеокласты - это симпластические макрофагальные структуры, образованные слиянием нескольких моноцитов крови. Межклеточное вещество состоит из основного аморфного вещества, импрегнированного неорганическими солями, в котором располагаются коллагеновые волокна, образующие небольшие пучки. Они содержат в основном белок- коллаген 1 и 5 типов. Волокна могут иметь беспорядочное или строго ориентированное направление. Из всех разновидностей соединительных тканей костная ткань обладает наиболее выраженными опорной, механической, защитной функциями для внутренних органов, а также является депо солей кальция, фосфора и др.Существует два основных стволовые полтипа костной ткани: ретикулофиброзная и пластинчатая.Эти разновидности костной ткани различаются по структурным и физическим свойствам. Возрастные изменения. Соединительные ткани с возрастом претерпевают изменения в строении, количестве и химическом составе, увеличиваются общая масса соединительнотканных образований, рост костного скелета. Во многих разновидностях соединительнотканных структур изменяется соотношение типов коллагена, гликозаминогликанов; в частности, в них становится больше сульфатированных соединений. 10. Кость как орган. Макро- и микроскопическое строение плоских и трубчатых костей. Кость (os) — это орган, являющийся компонентом системы органов опоры и движения, имеющий типичную форму и строение, характерную архитектонику сосудов и нервов, построенный преимущественно из костной ткани, покрытый снаружи надкостницей (periosteum) и содержащий внутри костный мозг (medulla osseum). Основу кости составляет компактное и губчатое вещества. Снаружи кость покрыта надкостницей. НАДКОСТНИЦА (периост)- плотная соединительная ткань, примыкающая к компактному в-ву кости. В ней остеобласты обеспечивающие рост кости в толщину.Кость строится из клеток, межклеточного вещества, коллагеновых волокон. Межклеточное в-во из тесно прилегающих костных пластинок, имеющих форму полых цилиндров разного диаметра, вставленных друг в друга и образующих остеоны.ОСТЕОН (гаверсова система)- структурная единица компактного в-ва кости, состоит из 5-20 костных пластинок, расположенных концентрически вокруг Гаверсова канала, что придаёт кости прочность, и образующая наружный слой эпифизов и диафиз кости.Через Гаверсов канал проходит 1 артерия, 1 вена, которые разветвляются на капилляры и подходят к лакунам данной гаверсовой системы, обеспечивая питание и дыхание остеобластов.Между костными пластинками лакуны, с остеобластами. На наружной и внутренней поверхности кости каналы Фолькмана с кровеносными сосудами, соединяющимися с сосудами Гаверсова канала. СТРОЕНИЕ ТРУБЧАТОЙ КОСТи: ГУБЧАТОЕ В-ВО- в-во кости состоящее из перемычек и балок(трабекул),образующих многочисленные ячейки.Трабекулы пересекаются в разных направлениях, их расположение соответствует направлению сил сжатия и растяжения, действующих на кость.Промежутки между трабекулами заполнены красным костным мозгом.Губчатое вещество находится в диафизах трубчатых костей, в коротких губчатых и плоских костях.ЭПИФИЗ- головка кости трубчатой. Заполнен губчатым в-вом в котором красный костный мозг.ДИАФИЗ- образует тело трубчатой кости из компактного в-ва. Внутри костномозговая полость с желтым костным мозгом 11. Характеристика основных этапов прямого остеогенеза. Способ остеогенеза характерен для развития грубоволокнистой ткани при образованииплоских костей, например покровных костей черепа.Процесс наблюдается в основном в течение первого месяца внутриутробного развития.В первой стадии-образование скелетогенного островка- в местах развития будущей кости происходят очаговое размножение мезенхимных клеток и васкуляризация скелетогенного островка. Во второй стадии, заключающейся в дифференцировке клеток островков, образуется оксифильное межклеточное вещество с коллагеновыми фибриллами- органическая матрица костной ткани. В основном веществе появляются мукопротеиды, цементирующие волокна в одну прочную массу. Некоторые клетки, дифференцируются в остеоциты, другие, располагающиеся по поверхности, дифференцируются в остеобласты. Остеобласты отделяются друг от друга. Постепенно эти клетки оказываются «замурованными» в межклеточном веществе, теряют способность размножаться и превращаются в остеоциты.Третья стадия-кальцификация межклеточного вещества. При этом остеобласты выделяют фермент щелочную фосфатазу, расщепляющую содержащиеся в периферической крови глицерофосфаты на углеводные соединения.Одним из посредников кальцификации является остеонектин-гликопротеин, избирательно связывающий соли кальция и фосфора с коллагеном. В результате кальцификации образуютсякостные перекладины, или балки. Затем от этих перекладин ответвляются выросты, соединяющиеся между собой и образующие широкую сеть. К моменту завершения гистогенеза по периферии зачатка кости в эмбриональной соединительной ткани появляется большое количество волокон и остеогенных клеток. Часть этой волокнистой ткани, прилегающей непосредственно к костным перекладинам, превращается впериост, который обеспечивает трофику и регенерацию кости. Далее в процессе развития она заменяется вторичной губчатой костью взрослых, которая отличается от первой тем, что построена из пластинчатой костной ткани- четвертая стадия остеогенеза.Костные пластинки образуются вокруг кровеносных сосудов путем дифференцировки, прилегающей к ним мезенхимы. Над такими пластинками образуется слой новых остеобластов и возникают новые пластинки. Таким образом, вокруг сосуда формируются костные цилиндры, вставленные один в другой. С момента появления остеонов ретикулофиброзная костная ткань перестает развиваться и заменяется пластинчатой костной тканью. Со стороны надкостницы формируются общие, или генеральные, пластинки, охватывающие всю кость снаружи. Так развиваются плоские кости. 12. Характеристика основных этапов непрямой остеогенеза. Непрямой остеогистогенез. Развитие кости на месте хряща,т.е. непрямой остеогенез,начинается в области диафиза.Образованию перихондриальной костной манжетки предшествует разрастание кровеносных сосудов с дифференцировкой в надхрящнице, прилежащей к средней части диафиза, остеобластов, образующих в виде манжетки сначала ретикулофиброзную костную ткань, затем заменяющуюся на пластинчатую.Образование костной манжетки нарушает питание хряща. Вследствие этого в центре диафизарной части хрящевого зачатка возникают дистрофические изменения. Хондроциты вакуолизируются, их ядра пикнотизируются, образуются пузырчатые хондроциты.Рост хряща в этом месте прекращается. Удлинение перихондральной костной манжетки сопровождается расширением зоны деструкции хряща и появлениемостеокластов-это приводит к появлению очагов эндохондрального окостенения. Таким образом, в колонке хондроцитов имею два противоположно направленных процесса- размножение и рост в дистальных отделах диафизы и дистрофические процессы в его проксимальном отделе.С момента разрастания сосудистой сети и появления остеобластов надхрящница превращается в надкостницу. Диафизарный хрящ разрушается, в нем возникают удлиненные пространства, в которых « поселяются» остеоциты, образующие на поверхности оставшихся участков обызвествленного хряща костную ткан VII. МЫШЕЧНЫ Е ТКАНИ 1. Морфофункциональная характеристика и гистогенетическая классификация мышечных тканей. Мышечные ткани представляют собой группу тканей различного происхождения и строения, но объединенных способностью к сокращению. Общая морфофункциональная характеристика мышечных тканей 1) структурные элементы (клетки, волокна) обладают удлиненной формой; 2) наличие органелл специального назначения – миофиламенты, миофибриллы; 3) с сократительными органеллами связаны элементы цитоскелета и плазмолемма; 4) расположение митохондрий рядом с сократительными элементами (обеспечение энергией в виде макроэргических соединений - АТФ); 5) наличие трофических включений гликогена, липидов, которые являются источником энергии; 6) наличие миоглобина – кислород-связывающего железосодержащего белка (в некоторых мышечных тканях); 7) хорошо развиты структуры, осуществляющие накопление и выделение ионов кальция (кавеолы, гладкая ЭПС); 8) для синхронизации сокращений соседние мышечные элементы иннервируются из одного источника или (и) связаны многочисленными щелевыми соединениями, которые обеспечивают транспорт ионов Классификация.В основу классификации мышечных тканей положены два принципа - морфофункциональный и гистогенетический. В соответствии с морфофункциональным принципом, в зависимости от структуры органелл сокращения, мышечные ткани подразделяют на две подгруппы. Первая подгруппа - поперечнополосатые (исчерченные) мышечные ткани (textus muscularis striatus). В цитоплазме их элементов миозиновые филамен-ты постоянно полимеризованы, образуют с актяновыми нитями постоянно существующие миофибриллы. Последние организованы в характерные комплексы - саркомеры. В соседних миофибриллах структурные субъединицы саркомеров расположены на одинаковом уровне и создают поперечную исчерченность. Исчерченные мышечные ткани сокращаются быстрее, чем гладкие. Вторая подгруппа - гладкие (неисчерченные) мышечные ткани (textus muscularis nonstriatus). Эти ткани характеризуются тем, что вне сокращения миозиновые филаменты деполимеризованы. В присутствии ионов кальция они полимеризуются и вступают во взаимодействие с филаментами актина. Образующиеся при этом миофибриллы не имеют поперечной исчерченности: при специальных окрасках они представлены равномерно окрашенными по всей длине (гладкими) нитями. В соответствии с гистогенетическим принципом в зависимости от источников развития (эмбриональных зачатков) мышечные ткани подразделяются на 5 типов: мезенхимные (из десмального зачатка в составе мезенхимы), эпидермальные (из кожной эктодермы и из прехордальной пластинки), нейральные (из нервной трубки), целомические (из миэпикардиальной пластинки висцерального листка сомита) и соматические (миотомные) 2. Морфофункциональная характеристика гладкой мышечной ткани. Гладкая мышечная ткань Этот тип мышечной ткани широко распространен в организме. Основные места ее локализации: 1) стенка сосудов 2) стенки внутренних органов пищеварительной, дыхательной, выделительной и половой систем 3) кожа (мышцы, поднимающие волос) 4) экзокринные железы (миоэпителиальные клетки) 5) радужная оболочка и цилиарное тело глаза. Строение гладкой мышечной ткани Структурно-функциональной единицей гладкой мышечной ткани является гладкий миоцит (ГМ) – веретеновидные, мало- и многоотростчатые (в эндокарде, аорте, мочевом пузыре). Большинство ГМ имеет веретеновидную форму длиной от 15 до 500 мкм и диаметром от 2 до 20 мкм. Ядро - сигарообразной формы, расположено вдоль длинной оси клетки в её центральной утолщенной части. При сокращении миоцита ядро образует складки и может штопорообразно закручиваться. Щелевые контакты связывают соседние ГМ и необходимы для проведения возбуждения (ионный ток), запускающего сокращение ГМ. В составе ГМ выделяют следующие структурно-функциональные аппараты: 1. трофико-энергетический аппарат – саркосомы, включения гликогена, липидов, миоглобина; обеспечивает энергией мышечное сокращение и другие энергоёмкие процессы, а также внутриклеточное депонирование и реализацию питательных веществ; 2. опорный аппарат – сарколемма, цитоскелет, плотные тельца (электронно-плотные структуры, расположенные в цитоплазме или связанные с сарколеммой, содержат белок α-актин) и межклеточные контакты (десмосомы, нексусы); обеспечивает формообразовательную, каркасную, локомоторную и интеграционную функции; 3. аппарат синтеза, структуризации и регенерации – свободные рибосомы, ЭПС, комплекс Гольджи, лизосомы; обеспечивает процессы внутриклеточной регенерации (в т.ч. ресинтез и сборку сократительных и регуляторных белков миофиламентов, а также секрецию компонентов базальной мембраны); 5. нервный аппарат – нервные волокна, чувствительные и двигательные нервные окончания вегетативной нервной системы; обеспечивает инициацию и регуляцию непроизвольного сокращения ГМ; 6. сократительный аппарат – актиновые (тонкие) и миозиновые (толстые) миофиламенты, способные к скольжению между собой; обеспечивает медленное длительное энергоёмкое кальций-зависимое сокращение-расслабление миоцита ("модель скользящих нитей"). 3. Морфофункциональная характеристика скелетной мышечной ткани. Структурные основы и механизм мышечного сокращения. Скелетные мышцы — активная часть опорно-двигательного аппарата, включающего также кости, связки, сухожилия и их сочленения. Развитие скелетных мышц. Источник развития – клетки миотомов сомитьов мезодермы, детерминированные в направлении миогенеза. Стадии: - миобласты - мышечные трубочки - дефинитивная форма миогенеза – мышечное волокно. Строение скелетной мышечной ткани. Структурно-функциональной единицей скелетной мышечной ткани является мышечное волокно. Оно представляет собой вытянутое цилиндрическое образование с заостренными концами, диаметром от 10 до 100 мкм, вариабельной длины (до 10-30 см.). Структурно-функциональной единицей скелетной мышечной ткани является мышечное волокно. Оно представляет собой вытянутое цилиндрическое образование с заостренными концами, диаметром от 10 до 100 мкм, вариабельной длины (до 10-30 см.). Механизм мышечного сокращенияописывается теорией скользящих нитей, которая была предложена Хью Хаксли. В покое, при очень низкой концентрации ионов Са++ в миофибрилле расслабленного волокна толстые и тонкие нити не соприкасаются. Толстые и тонкие филаменты беспрепятственно скользят относительно друг друга, в результате мышечные волокна не сопротивляются пассивному растяжению. Такое состояние свойственно мышце-разгибателю при сокращении соответствующего сгибателя. Мышечное сокращение вызывается резким повышением концентрации ионов Са++ и состоит из нескольких этапов: - Ионы Са++ связыватся с молекулой тропонина, которая смещается, открывая на тонких нитях участки связывания миозина. - Головка миозина прикрепляется к миозин-связывающим участкам тонкой нити. - Головка миозина изменяет конформацию и совершает гребковое движение, продвигающее тонкую нить к центру саркомера. - Головка миозина связывается с молекулой АТФ, что приводит к отделению миозина от актина. 4. Морфофункциональная характеристика сердечной мышечной ткани, типичных сократительных, проводящих и секреторных кардиомиоцитов. Сердечная мышечная ткань – поперечнополосатая мышечная ткань целомического типа. Она образует мышечную оболочку сердца, составляя 70-90% его массы. Источником развития сердечной мышечной ткани является миоэпикардиальная пластинка висцерального листка спланхнотома. Из нее выделяются миобласты, которые активно размножаются митозом и постепенно образуют миофибриллы. Структурно-функциональной единицей сердечной мышечной ткани является клетка – кардиомиоциты. По строению и функциям кардиомиоциты подразделяются на три основные группы: - типичные, или сократительные - атипичные, образующие проводящую систему - секреторные. Кардиомиоциты – цилиндрические или ветвящиеся клетки, длиной – 50-120 мкм (более крупные – желудочках, шириной –15-20 мкм. Снаружи покрыты сарколеммой. Атипичные кардиомиоциты.Обладают способностью к генерации и проведению электрических импульсов. Они образуют узлы и проводящую систему сердца. Характеризуются слабым развитием сократительного аппарата, светлой саркоплазмой и крупными ядрами. Не имеют Т-трубочек и не образуют вставочных дисков. Секреторные (эндокринные) кардиомиоциты- располагаются в предсердиях, характеризуются слабым развитием миофибрилл. Вырабатывают предсердный натрийуретический фактор. Он усиливает потерю натрия и воды из орзанизма, расширяет сосуды, уменьшает АД. 5. Закономерности эмбрионального миогенеза. Особенности регенерации различных видов мышечных тканей (А. Н. Студитский, А. А. Клишов). В мышечной, как в других тканях, различают два вида регенерации — физиологическую и репаративную. Физиологическая регенерация проявляется в форме гипертрофии мышечных волокон, что выражается в увеличении их толщины и даже длины, увеличение числа органелл, главным образом миофибрилл, а также нарастании числа ядер, что в конечном счете проявляется увеличением функциональной способности мышечного волокна. Радиоизотопным методом установлено, что увеличение числа ядер в мышечных волокнах в условиях гипертрофии достигается за счет деления клеток миосателлитов и последующего вхождения в миосимпласт дочерних клеток. Советским ученым А. Н. Студитским доказана возможность аутотрансплантации скелетной мышечной ткани и даже целых мышц при соблюдении определенных условий: механическое измельчение мышечной ткани трансплантата, с целью растормаживания клеток-сателлитов и последующей их пролиферации; помещение измельченной ткани в фасциальное ложе; подшивание двигательного нервного волокна к измельченному трансплантату; наличие сократительных движений мышц-антагонистов и синергистов. VIII. НЕРВНА Я ТКАНЬ 1. Морфофункциональная характеристика нервной ткани. Нервная ткань – это система взаимосвязанных нервных клеток и нейроглии, обеспечивающих специфические функции восприятия раздражений, возбуждения, выработки импульса и его передачи. Она является основой строения органов нервной системы, обеспечивающих регуляцию всех тканей и органов, их интеграцию в организме и связь с окружающей средой. Нервная ткань является функционально ведущей тканью НС и состоит из нейроцитов (нервных клеток) и нейроглии. Нейроны (нейроциты) обладают 4 свойствами: 1) способны воспринимать раздражение; 2) способны возбуждаться; 3) способны вырабатывать импульс; 4) способны передавать вырабатываемый импульс другим нейронам или на рабочие органы. Источник развития - дорсальная эктодерма. Нервная ткань развивается из нервного гребня, нейральных плакод и нервной трубки. 2. Нейроциты. Классификация. Особенности ультраструктурной организации. Свойства, проведение нервного импульса. Нейроны, или нейроциты – специализированные клетки нервной системы, ответственные за рецепцию, обработку (процессинг) стимулов, проведение импульса и влияние на другие нейроны, мышечные или секреторные клетки. Нейроны выделяют нейромедиаторы и другие вещества, передающие информацию. Нейрон является морфологически и функционально самостоятельной единицей. Нейроны имеют размеры от 4 до 140 мкм в диаметре, различную форму (пирамидную, звездчатую, паукообразную, круглую и др.). В то же время все нейроны имеют отростки длиной от нескольких микрометров до 1,5 м. Отростки подразделяются на 2 типа: 1) дендриты, которые ветвятся; их в нейроне может быть несколько, часто они короче аксонов; по ним импульс движется к телу клетки; 2) аксоны, или нейриты; нейрит в клетке может быть только 1; по аксону импульс движется от тела клетки и передается на рабочий орган или на другой нейрон. Морфологическая классификация нейроцитов (по количеству отростков). В зависимости от количества отростков нейроциты подразделяются на: 1) униполярные, если имеется только 1 отросток (аксон); встречаются только в эмбриональном периоде; 2) биполярные, содержат 2 отростка (аксон и дендрит); встречаются в сетчатке глаза и спиральном ганглии внутреннего уха; 3) мультиполярные — имеют более 2 отростков, один из них — аксон, остальные — дендриты; встречаются в головном и спинном мозге и периферических ганглиях вегетативной нервной системы; 4) псевдоуниполярные — это фактически биполярные нейроны, так как аксон и дендрит отходят от тела клетки в виде одного общего отростка и только потом разделяются и идут в различных направлениях; находятся в чувствительных нервных ганглиях (спинномозговых, чувствительных ганглиях головы). По функциональной классификации нейроциты подразделяются на: 1) чувствительные, их дендриты заканчиваются рецепторами (чувствительными нервными окончаниями); 2) эффекторные, их аксоны заканчиваются эффекторными (двигательными или секреторными) окончаниями; 3) ассоциативные (вставочные), соединяют друг с другом два нейрона. 3. Нейроглия. Источники происхождения, классификация, функции. Нейроглия (греческое neuron – нерв, glia – клей) – термин, введенный для описания связующих элементов между нейронами. Глиоциты – разнообразные вспомогательные клетки нервных тканей. Это обширная гетерогенная группа элементов нервной ткани,обеспечивающая деятельность нейронов и выполняющая широкий круг функций. Функции нейроглии: 1. Опорная. 2. Трофическая. 3. Разграничительная. 4. Поддержание постоянства среды вокруг нейронов. 5. Секреторная. 6. Защитная. Классификация нейроглии. Нейроглия включает макроглию и микроглию. Макроглия подразделяется на: 1. Эпендимная глия – образована клетками кубической или цилиндрической формы, однослойные пласты которых выстилают полости желудочков головного мозга и центрального канала спинногомозга. 2. Олигодендроглия – то есть глия с малым количеством отростков. Это обширная группа разнообразных мелких клеток, с короткими немногочисленными отростками, которые окружают тела ней-ронов, входят в состав нервных волокон и нервных окончаний. 3. Астроглия – представлена астроцитами – самыми крупными глиальными клетками. Микроглия – представляет собой совокупность мелких удли-ненных звездчатых клеток с плотной цитоплазмой и сравнительно короткими ветвящимися отростками. Она располагается преимуще ственно вдоль капилляров в центральной нервной системе. Имеет мезенхимное происхождение. По морфологии выделяют несколько типов микроглии: 1. Покоящаяся (типичная, ветвистая) 2. Амебоидная – временная форма микроглии, обнаружена в развивающемся мозге. 3. Реактивная – появляется после травмы, не имеет ветвящихся отростков, не имеет филоподий и псевдоподий. 4. Нервные волокна миелиновые и безмиелиновые. Нерв как орган. Особенности регенерации нервных волокон. Отростки нервных клеток, покрытые оболочками, называются нервными волокнами. По строению оболочек различают миелиновые и безмиелиновые нервные волокна Безмиелиновые нервные волокна находятся преимущественно в составе автономной, или вегетативной, нервной системы. Нейролеммоциты оболочек безмиелиновых нервных волокон, располагаясь плотно, образуют тяжи. Такие волокна, содержащие несколько осевых цилиндров, называются волокнами кабельного типа. По мере погружения осевых цилиндров в тяж нейролеммоцитов оболочки последних прогибаются, плотно охватывают осевые цилиндры и, смыкаясь над ними, образуют глубокие складки, на дне которых и располагаются отдельные осевые цилиндры. Сближенные в области складки участки оболочки нейролеммоцита образуют сдвоенную мембрану — мезаксон, на которой как бы подвешен осевой цилиндр. Миелиновые нервные волокна встречаются как в центральной, так и в периферической нервной системе. Встречаются в ЦНС и ПС. ● Скорость проведения нервного импульса выше, чем в безмиел новых нервных волокнах. ● Волокна толще. ● Содержат один осевой цилиндр. ● В составе оболочки имеется миелиновый слой (спирально закрученный длинный мезаксон). ● Встречаются насечки миелина (насечки Шмидта –Лантермана). ● Имеются узловые перехв Регенерация зависит от места травмы. Как в центральной, так и в периферической нервной системе погибшие нейроны не восстанавливаются. Полноценной регенерации нервных волокон в центральной нервной системе обычно не происходит, но нервные волокна в составе периферических нервов обычно хорошо регенерируют. 5. Нервные окончания: понятие, классификация. Строение рецепторных окончаний. Нервные окончания – это концевые аппараты, которыми заканчиваются нервные волокна. По функциям все нервные окончания разделяются на три группы: Межнейрональные (синапсы) – обеспечивают функциональную связь между нейронами. Эффекторные (двигательные) – передают сигналы из ЦНС на исполняющие органы (мышцы, железы). Располагаются на терменалях аксона. Рецепторные (чувствительные) – воспринимают раздражения из внешней и внутренней среды, находятся на дендритах. Рецепторные нервные окончания. Эти нервные окончания — рецепторы — рассеяны по всему организму и воспринимают различные раздражения как из внешней среды, так и от внутренних органов. Соответственно выделяют две большие группы рецепторов: экстерорецепторы и интерорецепторы. К экстерорецепторам (внешним) относятся: слуховые, зрительные, обонятельные, вкусовые и осязательные рецепторы. К интерорецепторам (внутренним) относятся: висцеро-рецепторы (сигнализирующие о состоянии внутренних органов) и проприорецепторы (или рецепторы опорно-двигательного аппарата). 6. Нервные окончания: понятие, классификация. Строение эффекторных окончаний. Нервные окончания – это концевые аппараты, которыми заканчиваются нервные волокна. По функциям все нервные окончания разделяются на три группы: Межнейрональные (синапсы) – обеспечивают функциональную связь между нейронами. Эффекторные (двигательные) – передают сигналы из ЦНС на исполняющие органы (мышцы, железы). Располагаются на терменалях аксона. Рецепторные (чувствительные) – воспринимают раздражения из внешней и внутренней среды, находятся на дендритах. Эффекторные нервные окончания. Среди эффекторных нервных окончаний различают двигательные и секреторные. Двигательные нервные окончания — это концевые аппараты аксонов двигательных клеток соматической или вегетативной нервной системы. При их участии нервный импульс передается на ткани рабочих органов. Двигательные окончания в поперечнополосатых мышцах называются нервно-мышечными окончаниями. Они представляют собой окончания аксонов клеток двигательных ядер передних рогов спинного мозга или моторных ядер головного мозга. Двигательные нервные окончания в гладкой мышечной ткани представляют собой чёткообразные утолщения (или варикозы) нервного волокна, идущего среди неисчерченных гладких миоцитов. Варикозы содержат адренергические или холинергические пресинаптические пузырьки. Нейролеммоциты в области варикозов часто отсутствуют, и волокно проходит «обнаженным». Сходное строение имеют секреторные нервные окончания (нейрожелезистые). Они представляют собой концевые утолщения терминали или утолщения по ходу нервного волокна, содержащие пресинаптические пузырьки, главным образом холинергические. 7. Синапсы. Синапс— специализированные структуры, которые обеспечивают передачу возбуждения с одной возбудимой клетки на другую. Понятие СИНАПС введено в физиологию Ч.Шеррингтоном (соединение, контакт). Синапс обеспечивает функциональную связь между отдельными клетками. Подразделяются на нервно-нервные, нервно-мышечные и синапсы нервных клеток с секреторными клетками (нервно-железистые). В нейроне выделяется три функциональных отдела: сома, дендрит, аксон. Поэтому между нейронами существуют все возможные комбинации контактов. Например, аксо-аксональный, аксо-соматический и аксо-дендритный. Классификация. 1)по местоположению и принадлежности соответствующим структурам: - периферические(нервномышечные, нейросекреторные, рецепторнонейрональные); - центральные(аксо-соматические, аксо-дендритные, аксоаксональные, сомато-дендритные. сомато-соматические); 2)механизму действия— возбуждающие и тормозящие; 3)способу передачи сигналов— химические, электрические, смешанные. 4)химические классифицируют по медиатору, с помощью которого осуществляется передача— холинергические, адренергические, серотонинергические, глицинергически. и т.д. Строение синапса. Синапс состоит из следующих основных элементов: •Пресинаптической мембраны (в нервномышечном синапсе — это концевая пластинка): •Постсинаптической мембраны; •Синаптической щели. Синаптическая щель заполнена олигосахаридсодержащей соединительной тканью, которая играет роль поддерживающей структуры для обеих контактирующих клеток. 8. Рефлекторные дуги. Нейронная теория, вклад зарубежных и отечественных ученых в ее становлении (С. Рамон-и-Кахал, К. Гольджи, А. С. Догель, Б. И. Лаврентьев). Рефлекторные дуги. Нервная ткань входит в состав нервной системы, функционирующей по рефлекторному принципу, морфологическим субстратом которого является рефлекторная дуга. Рефлекторная дуга представляет собой цепь нейронов, связанных друг с другом синапсами и обеспечивающих проведение нервного импульса от рецептора чувствительного нейрона до эфферентного окончания в рабочем органе. Самая простая рефлекторная дуга состоит из двух нейронов — чувствительного и двигательного. В подавляющем большинстве случаев между чувствительными и двигательными нейронами включены вставочные, или ассоциативные, нейроны. У высших животных рефлекторные дуги состоят обычно из многих нейронов и имеют значительно более сложное строение. Нейронная теория. Теория контакта, утверждающая, что нервная система построена из обособленных, контактирующих между собой клеток - нейронов, сохраняющих генетическую, морфологическую и функциональную индивидуальность. Н. т. рассматривает нервную деятельность как результат взаимодействия совокупности нейронов. Этому представлению в конце 19 - начале 20 вв. противостояла теория континуитета, полагавшая, что клеточное вещество одного нейрона переходит в вещество другого без перерыва, благодаря чему отростки нервных клеток образуют единую плазматическую сеть. Сторонники этой теории (венгерский учёный И. Апати, немецкий - А. Бете и др.) считали, что цитоплазматическую непрерывность нервной ткани обеспечивают нейрофибриллы. Убедительные факты в пользу Н. т. были получены С. Рамон-и-Кахалем, А. А. Заварзиным, Б. И. Лаврентьевым и др. при изучении микроскопического строения нервной системы, её эмбрионального развития, а также дегенерации и регенерации нейронов. Ныне в свете электрофизиологических и электронномикроскопических данных правильность Н. т. не вызывает сомнений. Нервная система у всех организмов, включая низшие, образована обособленными нейронами, взаимодействующими в местах контакта, которые имеют сложное строение и называются синапсами. Отступления от этого общего принципа редки. Функциональная обособленность нейронов может утрачиваться при синхронном возбуждении группы нейронов (например, в центре, иннервирующем электрические органы рыб). У кальмаров наличие гигантских аксонов объясняется плазматическим слиянием отростков нескольких нейронов, утративших морфологическую обособленность. 9. Нервные окончания в эпителиальных тканях. Нервные волокна заканчиваются концевыми аппаратами — нервными окончаниями. Различают 3 группы нервных окончаний: концевые аппараты, образующие межнейрональные синапсы и осуществляющие связь нейронов между собой; эффекторные окончания (эффекторы), передающие нервный импульс на ткани рабочего органа; рецепторные (аффекторные, или чувствительные). По особенностям строения чувствительные окончания делятся на свободные нервные окончания, в составе которых имеются только конечные разветвления осевого цилиндра, и несвободные нервные окончания, в состав которых входят все компоненты нервного волокна (разветвление осевого цилиндра, клетки глии). Рецепторные нервные окончания в составе эпителиальной, соединительной и мышечной тканей имеют целый ряд особенностей. В эпителиальной ткани локализованы свободные нервные окончания. При их формировании миелиновые нервные волокна, подойдя к эпителиальному пласту теряют миелин, а осевые цилиндры проникают в эпителий и распадаются в нем между клетками на тонкие терминальные ветви. Функция свободных рецепторов, например, эпидермиса, связана с восприятием болевых и температурных раздражений. Свободные нервные окончания могут в виде корзинки охватывать волосяные фолликулы. Регистрируя перемещения в пространстве, отдельные волоски, играют роль механорецепторов. 10. Нервные окончания в соединительных тканях. Нервные волокна заканчиваются концевыми аппаратами — нервными окончаниями. Различают 3 группы нервных окончаний: концевые аппараты, образующие межнейрональные синапсы и осуществляющие связь нейронов между собой; эффекторные окончания (эффекторы), передающие нервный импульс на ткани рабочего органа; рецепторные (аффекторные, или чувствительные). По особенностям строения чувствительные окончания делятся на свободные нервные окончания, в составе которых имеются только конечные разветвления осевого цилиндра, и несвободные нервные окончания, в состав которых входят все компоненты нервного волокна (разветвление осевого цилиндра, клетки глии). Весьма разнообразны рецепторы в соединительной ткани и соответственно делятся на несвободные некапсулированные, инкапсулированные и нервно-сухожильные веретена. Инкапсулированные рецепторы соединительной ткани при всем их разнообразии всегда состоят из разветвленного осевого цилиндра и глиальных клеток, которые снаружи покрыты соединительнотканной капсулой. Среди инкапсулированных нервных окончаний в соединительной ткани в зависимости от строения различают пластинчатые тельца Фатер-Пачини, луковицевидные тельца Гольджи-Маццони, осязательные тельца Мейснера, конечные колбы Краузе. 11. Нервные окончания в мышечных тканях. Нервные волокна заканчиваются концевыми аппаратами — нервными окончаниями. Различают 3 группы нервных окончаний: концевые аппараты, образующие межнейрональные синапсы и осуществляющие связь нейронов между собой; эффекторные окончания (эффекторы), передающие нервный импульс на ткани рабочего органа; рецепторные (аффекторные, или чувствительные). Моторные нервные окончания на скелетной мышечной ткани называются нервно-мышечными окончаниями, или моторными бляшками (terminatio neuromuscularis). Моторные бляшки образуются следующим образом. При подходе нервного волокна к мышечному, нервное волокно утрачивает миелиновый слой оболочки, неврилемма сливается с базальной мембраной сарколеммы, а освобожденные от оболочки терминали погружаются в глубь саркоплазмы, увлекая за собой плазмолемму сарколеммы, которая, как перчатка пальцы, одевает каждую терминалы. В результате образуются 2 полюса нервно-мышечного окончания: нервный и мышечный. Частная гистология 1Х. Нервная система 1. Нерв. Строение, тканевой состав. Реакция на повреждение, регенерация. Нервы (нервные стволы) – это структуры, которые связывают центры головного и спинного мозга с рецепторами и рабочими органами. Они состоят из миелиновых и безмиелиновых афферентных и эфферентных волокон и прослоек соединительной ткани. Могут быть отдельные нейроны и отдельные нервные ганглии. Прослойка рыхлой соединительной ткани, окружающая каждое нервное волокно, называется эндоневрий; окружающая пучок нервных волокон —периневрий, который состоит из 5-6 слоев коллагеновых волокон; между этими слоями имеются щелевидные полости, выстланные нейроэпителием, в которых циркулирует жидкость. Весь нерв окружен прослойкой соединительной ткани, которая называется эпиневрий. В периневрии и эпиневрии имеются кровеносные сосуды и нервы нервов. Регенерация. При повреждении нерва разрываются проходящие в нем нервные волокна. После разрыва волокна в нем образуются 2 конца — конец, который связан с телом нейрона, называется центральным; конец, не связанный с нервной клеткой, называется периферическим.В периферическом конце происходят 2 процесса: 1) дегенерация и 2) регенерация. Вначале идет процесс дегенерации, заключающийся в том, что начинается набухание нейролеммоцитов, растворяется миелиНовый слой, осевой цилиндр фрагментируется, образуются капли (овоиды), состоящие из миелина и фрагмента осевого цилиндра. К концу 2-й недели происходит рассасывание овоидов, остается только неврилемма оболочки волокна. Нейролеммоциты продолжают размножаться, из них образуются ленты (тяжи). После рассасывания овоидов осевой цилиндр центрального конца утолщается и образуется колба роста, которая начи- нает расти, скользя по лентам нейролеммоцитов. К этому времени между разорванными концами нервных волокон образуется нейроглиально-соединительнотканный рубец, являющийся препятствием для продвижения колбы роста. Поэтому не все осевые цилиндры могут пройти на противоположную сторону образовавшегося рубца. Следовательно, после повреждения нервов иннервация органов или тканей полностью не восстанавливается. Между тем часть осевых цилиндров, оснащенных колбами роста, пробивается на противоположную сторону нейроглиального рубца, погружается в тяжи нейролеммоцитов. Затем мезаксон навертывается на эти осевые цилиндры, образуется миелиновый слой оболочки нервного волокна. В том месте, где находится нервное окончание, рост осевого цилиндра приостанавливается, формируются терминали окончания и все его компоненты. 2. Чувствительные нервные узлы (спинномозговые и черепные). Строение, тканевой состав. Цитофункциональная характеристика нейронов и нейроглии. ЧУВСТВИТЕЛЬНЫЕ УЗЛЫ Чувствительные узлы лежат по ходу задних корешков спинного мозга либо черепномозговых нервов.Спинномозговой узел окружен соединительнотканной капсулой. От капсулы в паренхиму узла проникают тонкие прослойки соединительной ткани, которая образует его остов и проводит кровеносные сосуды. Нервные узлы могут быть чувствительными и вегетативными. Нейроны узла могут быть псевдоуниполярными (спинномозговой узел) и мультиполярными (вегетативные нервные узлы). Нейроны, образующие спинномозговой узел, располагаются группами на его периферии. Нейроны спинномозговых узлов чувствительные, а вегетативных эфферентные. Нервные клетки спинномозговых узлов окружены слоем клеток глии, которые здесь называются мантийными глиоцитами, или глиоцитами ганглия. Они расположены вокруг тела нейрона и имеют округлые ядра. Снаружи глиальная оболочка тела нейрона покрыта тонковолокнистой соединительнотканной оболочкой. Клетки этой оболочки отличаются овальной формой ядер. 3. Спинной мозг. Общая характеристика строения. Строение серого вещества: виды нейронов и их участие в образовании рефлекторных дуг, типы глиоцитов. Ядра серого вещества. Строение белого вещества.Спинной мозг состоит из серого вещества, расположенного центрально, вокруг спинномозгового канала, заполненного спинномозговой жидкостью, и окружающего его белого вещества. Серое вещество состоит из расположенных группами мультиполярных нейронов, нейроглиоцитов, безмиелиновых и тонких миелиновых волокон. Скопления нейронов, имеющих общую морфологию и функцию, называются ядрами. Серое вещество спинного мозга состоит из тел нейронов, безмиелиновых и тонких миелиновых волокон и нейроглии. Составной частью, являются мультиполярные нейроны. В сером веществе спинного мозга различают 3 типа клеток:1. Внутренние – их отростки заканчиваются в пределах серого вещества своей половины или противоположной стороны.2. Пучковые – их отростки выходят в белое вещество и образуют пучки проводящих волокон (восходящих и нисходящих), связывают между собой отдельные сегменты спинного мозга или поднимаются в продолговатый мозг.3. Корешковые – это аксоны мотонейронов передних рогов спинного мозга, образующие передние корешки. Белое вещество представляет собой совокупность продольно ориентированных преимущественно миелиновых волокон. Пучки нервных волокон, осуществляющие связь между различными отделами нервной системы, называются проводящими путями спинного мозга. Отдельные участки серого вещества спинного мозга значительно отличаются друг от друга по составу нейронов, нервных волокон и нейроглии. В задних рогах различают губчатый слой, желатинозное вещество, собственное ядро заднего рога и грудное ядро. Между задними и боковыми рогами серое вещество вдаётся тяжами в белое, вследствие чего образуется сетеобразное разрыхление, получившее название сетчатого образования. Рефлекторная дуга (нервная дуга) — путь, проходимый нервными импульсами при осуществлении рефлекса. Рефлекторная дуга состоит из:рецептора — нервное звено, воспринимающее раздражение;афферентного звена — центростремительное нервное волокно —отростки рецепторных нейронов, осуществляющие передачу импульсов от чувствительных нервных окончаний в центральную нервную систему;центрального звена — нервный центр (необязательный элемент, например для аксон-рефлекса);эфферентного звена — осуществляют передачу от нервного центра кэффектору.эффектора — исполнительный орган, деятельность которого изменяется в результате рефлекса.Различают: — моносинаптические, двухнейронные рефлекторные дуги;— полисинаптические рефлекторные дуги (включают три и более нейронов).Простейшая рефлекторная дуга у человека образована двумя нейронами — сенсорным и двигательным (мотонейрон) Белое вещество располагается снаружи от серого и делится серым веществом на три канатика - передний, задний и боковой. Вбелом веществе проходят толстые миелиновые нервные волокна, которые образуют проводящие пути спинного мозга – короткие, восходящие и нисходящие пучки.Короткие пучки ассоциативных волокон связывают сегменты спинного мозга, расположенные на различных уровнях.Восходящие (афферентные, чувствительные) пучки направляются к центрам головного мозга и мозжечка.Нисходящие (эфферентные, двигательные) пучки идут от головного мозга к клеткам передних рогов спинного мозга.В белом веществе передних канатиков находятся преимущественно нисходящие проводящие пути, в боковых канатиках – восходящие и нисходящие проводящие пути, в задних канатиках – восходящие проводящие пути.На поперечном срезе, окрашенном AgNO3, миелиновые волокна имеют вид кружков – внутри темная точка - осевой цилиндр и пустое пространство по периферии - цитолемма швановской клетки.Глия. Центральный канал изнутри выстлан эпиндимной глией, участвующей в секреции и перемещении спинномозговой жидкости. Базальный отросток этих клеток участвует в образовании наружной пограничной мембраны. В спинном мозге располагаются протоплазматические и волокнистые астроциты. Отростки волокнистых астроцитов, выходят в белое вещество и образуют там глиальные перегородки, отделяя канатики и другие структуры. Кроме того, в сером веществе находятся олигодендроглия – образует оболочки нервных волокон и микроглия. 3. Головной мозг (большие полушария). Цитоархитектоника слоев коры больших полушарий, нейронный состав. Представление о модульной организации коры. Миелоархитектоника – радиальные и тангенциальные нервные волокна. Гемато-энцефалический барьер, его строение и функции. Кора больших полушарий имеет различную толщину от 3 до 5 мм, формирующая борозды и извилины. Снаружи покрыта мягкой мозговой оболочкой, содержащей сосуды и нервы, которые обеспечивают трофику коры. Кора больших полушарий и мозжечок, представляют собой экранные нервные центры, клетки в них располагаются слоями в виде экрана. Цитоархитектоника коры полушарий головного мозга.Сразу под мягкой мозговой оболочкой располагается:1. Молекулярный слой – в нем находится небольшое количество веретеновидных клеток, а также тангенсальное сплетение, расположенное параллельно поверхности и образованное волокнами, как молекулярного слоя, так и нижележащих слоев коры полушарий.2. Наружный зернистый слой– он содержит большое количество густо расположенных клеток. Клетки этого слоя имеют различную форму (звездчатую, паукообразную, угловатую и др.), их отростки образуют связи между соседними клетками, а также поднимаются в молекулярный слой и принимают участие в формировании тангенсального сплетения. 3. Пирамидный слой– представлен клетками пирамидной формы, размером от 10 до 40 мкм. От вершины отходит главный дендритит, который поднимаются в молекулярный слой, и принимает участие в формировании тангенсального сплетения, от боковых поверхностей этих клеток отходят дендриты, обеспечивающие связь с соседними клетками. Аксоны этих клеток отходят от середины их основания и образуют ассоциативные нервные волокна, соединяющие клетки между собой в одном полушарии и коммисуральные нервные волокна, проходящие в мозолистом теле и связывающие клетки с соседним полушарием.4. Внутренний зернистый слой– он имеет подобное строение как наружный зернистый слой и содержит множество мелких нейронов, густо расположенных, но в данном слое преобладают клетки звездчатой формы. Отростки этого слоя также поднимаются в молекулярный слой и принимают участие в образовании тангенсального сплетения.5. Ганглионарный слой (слой больших пирамид)– он представлен крупными нейронами, которые были описаны В.А. Бецом и получили название – клетки Беца. По строению подобны малым пирамидам, но их размер 80 на 120 мкм. Дендриты, отходящие от вершины участвуют в образовании тангенсального сплетения молекулярного слоя, а от боковых поверхностей клеток обеспечивают связь с соседними нейронами. Аксоны, отходящие от середины основания клетки, образуют проекционные (двигательные, эфферентные) нервные волокна, формирующие нисходящие проводящие пути.6. Полиморфный слой– он образован клетками различной формы, но с преобладанием веретеновидных. В процессе закладки более крупные клетки расположены на границе с белым веществом, а клетки помельче прилежат к ганглионарному слою. Отростки данного слоя, также достигают молекулярного слоя и принимают участие в формировании тангенсального сплетения. Миелоархитектоника коры больших полушарии головного мозгапредставлена следующими структурами:1.Тангенсальное сплетение молекулярного слоя.2.Внешние и внутренние полосы Белларже, расположенные между внутренним зернистым и ганглионарным слоями, представляет собой тангенсальное сплетение, которое обеспечивает горизонтальные связи между клетками.3.Ассоциативные волокна, образованные аксонами пирамидных клеток.4.Коммисуралные волокна, образованные аксонами пирамидных клеток.5.Проекционные волокна, образованные аксонами клеток Беца.Различают 2 вида коры:· Гранулярный тип – при данном типе в коре будут хорошо развиты наружный и внутренний зернистые слои и плохо развиты пирамидный слой (височная, затылочная доли). Хорошо развиты II и IV слои коры больших полушарий.· Агранулярный тип – плохо развиты зернистые слои, но хорошо пирамидный, ганглионарнй и полиморфный слои коры (прецентральная извилина). Хорошо развиты III, V и VI слои коры больших полушарий.МОДУЛЬ.Модуль – это структурно-функциональная единица неокортекса, описанная школой Сентаготаи. Возбуждение в коре передается в виде вертикальных колонок, через все слои коры. Модуль имеет диаметр около 300 мкм и состоит из 2 микромодулей (Д до 100 мкм). Модуль проецируется а 3 модуля свого полушария и 2 противоположного. В состав модуля входят: Пирамидные клетки,3 афферентных волокна – 1 кортикокортикальное и 2 таламокортикальных. Возбуждающие нейроны – шипиковые нейроны, на их отростках находятся шипики:o диффузногоo фокального типа.Тормозные нейроны:o Корзинчатые (мелкие и крупные),o Аксо-аксональные нейроны,o Нейроны с аксональной кисточкой,o Нейроны с двойным букетом дендритов – это нейрон тормозит тормозные нейроны, тем самым приводит к возбуждению пирамидных нейронов (вызывает вторичное возбуждение пирамидных клеток)Т.о., в модуле есть 3 отдела:1) вход – таламокортикальные и кортикокортикальное волокна, несущие импульс от таламуса,2) зона обработки информации – система пирамидных и звездчатых клеток, связанных активирующими и тормозными синапсами,3) выход – аксоны пирамидных клеток. Гематоэнцефалический барьер регулирует проникновение из крови в мозг биологически активных веществ, метаболитов, химических веществ, воздействующих на чувствительные структуры мозга, препятствует поступлению в мозг чужеродных веществ, микроорганизмов, токсинов.В представлениях о гематоэнцефалическом барьере в качестве основных положений подчеркивается следующее:1) проникновение веществ в мозг осуществляется главным образом не через ликворные пути, а через кровеносную систему на уровне капилляр — нервная клетка;2) гематоэнцефалический барьер является в большей степени не анатомическим образованием, а функциональным понятием, характеризующим определенный физиологический механизм. Как любой существующий в организме физиологический механизм, гематоэнцефалический барьер находится под регулирующим влиянием нервной и гуморальной систем;3) среди управляющих гематоэнцефалическим барьером факторов ведущим является уровень деятельности и метаболизма нервной ткани. 4. Мозжечок. Строение и нейронный состав коры мозжечка. Межнейрональные связи. Афферентные и эфферентные нервные волокна. Глиоциты мозжечка. МОЗЖЕЧОК.Представляет собой центральный орган равновесия и координации движений. Состоит из 2 симметричных половин, соединенных червем. Содержит белое вещество, расположенное в центре, по периферии серое вещество – кора мозжечка, образующая борозды и извилины. Борозды и извилины создают на разрезе характерную для мозжечка картину «древа жизни». Снаружи мозжечок покрыт мягкой мозговой оболочкой с сосудами и нервами.В коре мозжечка выделяют 3 слоя: молекулярный слой, ганглионарный слой, зернистый слой. Ганглионарный слой – образован клетками грушевидной формы или клетками Пуркинье, лежащими в 1 слой. Это эфферентные нейроны. Тело этих клеток грушевидной формы, от тела в молекулярный слой отходят 2-3 дендрита, которые образуют густые ветвления в одной плоскости, в плоскости поперек извилины в виде шпалер и в другой плоскости - Молекулярный слой – он содержит 2 вида клеток:1.Корзинчатые клетки – они лежат в нижней трети молекулярного слоя, имеют длинные ветвящиеся дендриты и длинный аксон, расположенный в плоскости поперек извилины. 2.Звездчатые клетки – выполняют вставочную функцию и обеспечивают торможение клеток Пуркинье.· Мелкие звездчатые клетки с короткими отростками, они образуют синаптическую связь сдендритами клеток Пуркинье.· Крупные звездчатые нейроны с длинными отростками – его дендриты образуют связи с дендритами грушевидных клеток (дендро-дендритическая сочетательная система мозжечка), а их аксон опускается к телу клетки Пуркинье и участвует в образовании корзинки.Зернистый слой – образован 4 видами клеток:1. Клетка-зерно – она имеет маленький перикарион (5-8 мкм). От тела отходят 3-4 дендрита, которые ветвятся в виде лапок птиц. 2. Большой звездчатый нейрон с короткими отростками – его аксон проксимальнее клубочков мозжечка контактирует с дендритами клеток-зерен, а дендрит образует связь с аксоном клетки-зерна..3. Большой звездчатый нейрон с длинными отростками – образует ассоциативные икоммисуральные нервные волокна мозжечка.4. Веретеновидная горизонтальная клетка – расположена на границе ганглионарного и зернистого слоев. Имеет вытянутое тело, ее отростки образуют горизонтальные ветвления, которые заканчиваются в ганглионарном слое и выходят в белое вещество. Глиоциты. Кора мозжечка содержит различные глиальные элементы. В зернистом слое имеются волокнистые и протоплазматические астроциты. Ножки отростков волокнистых астроцитов образуют периваскулярные мембраны.Во всех слоях в мозжечке имеются олигодендроциты. Особенно богаты этими клетками зернистый слой и белое вещество мозжечка. В ганглионарном слое между грушевидными нейронами лежат глиальные клетки с темными ядрами. Отростки этих клеток направляются к поверхности коры и образуют глиальные волокна молекулярного слоя мозжечка. Афферентное и эфферентное нервные волокна коры мозжечка Афферентные волокна, поступающие в кору мозжечка, представлены двумя видами — моховидными и так называемыми лазящими волокнами. Моховидные волокна идут в составе оливомозжечкового и мостомозжечкового путей и опосредованно через клеткизерна оказывают на грушевидные клетки возбуждающее действие. Лазящие волокна поступают в кору мозжечка, по-видимому, по спинно-мозжечковому и вестибуломозжечковому путям. Они пересекают зернистый слой, прилегают к грушевидным нейронам и стелются по их дендритам, заканчиваясь на их поверхности синапсами. Лазящие волокна передают возбуждение непосредственно грушевидным нейронам. 5. Автономная (вегетативная) нервная система. Общая характеристика строения центральных и периферических отделов симпатической и парасимпатической систем. Строение и нейронный состав ганглиев (экстрамуральных и интрамуральных). Пре- и постганглионарные нервные волокна. Вегетативные ганглии представляют собой скопление многочислен-ных мультиполярных нервных клеток.Величина вегетативных ганглиев существенно варьирует. В связи с этим различают крупные, средней величины, мелкие и очень мелкие (микроганглии) ганглии. Вегетативный ганглий с поверхности покрыт фиброзной соединительнотканной капсулой, от которой внутрь отходят многочисленные прослойки соединительной ткани, образующей строму узла. По этим прослойкам в узел проходят кровеносные сосуды, питающие его и образующие в нем капиллярную сеть. В капсуле и строме узла часто вблизи кровеносных сосудов встречаются рецепторы – диффузные, кустиковидные или инкапсулированные. Симпатический отдел по своим основным функциям является трофическим. Он обеспечивает усиление окислительных процессов, усиление дыхания, учащение деятельности сердца, Парасимпатический отдел выполняет охранительную роль (сужение зрачка, бронхов, снижение частоты сердечных сокращений, опорожнение полостных органов), ее тонус преобладает ночью («царство вагуса»). Симпатический и парасимпатический отделы отличаются также и медиаторами - веществами, осуществляющими передачу нервных импульсов в синапсах. Медиатором в симпатических нервных окончаниях является норадреналин. Медиатором парасимпатических нервных окончаний - ацетилхолин. Наряду с функциональными имеются ряд морфологических различий симпатического и парасимпатического отделов вегетативной нервной системы, а именно: Парасимпатические центры разобщены, находятся в трех отделах мозга (мезенцефалический, бульбарный, сакральный), а симпатические - в одном (тораколюмбальный отдел). К симпатическим узлам относятся узлы I и II порядка, к парасимпатическим - III порядка (конечные). В связи с чем преганглионарные симпатические волокна более короткие, а постганглионарные более длинные, чем парасимпатические. Парасимпатический отдел имеет более ограниченную область иннервации, иннервируя только внутренние органы. Симпатический же отдел иннервирует все органы и ткани. Автономная (вегетативная) система из нервной системы выделена в связи с особенностями ее строения и функций. У всей нервной системы, исходя из ее функций, выделяют соматическую ее часть и автономную (вегетативную). К соматической части относят те отделы нервной системы, которые участвуют в выполнении функций, подконтрольных нашему сознанию. Это структуры, с помощью которых организм устанавливает взаимоотношения с внешней средой, воспринимает различные виды чувствительности — болевую, температурную, тактильную, световую, звуковую, вкусовую, а также обеспечивает функции опорно-двигательного аппарата. Вегетативная часть нервной системы осуществляет иннервацию всех обменных процессов в теле человека, поддерживает постоянство внутренней среды, координирует функции внутренних органов, желез, сердечно-сосудистой системы. Х. СЕРДЕЧНО — СОСУДИСТАЯ СИСТЕМА 1. Кровеносные сосуды. Общий принцип строения, тканевой состав, классификация. Сердечно – сосудистая система включает сердце, кровеносные и лимфатические сосуды. При этом сердце, кровеносные и лимфатические сосуды называются системой кровообращения или кровеносной системой. Лимфатические сосуды вместе с лимфатическими узлами относятся к лимфатической системе. Кровеносная система – это замкнутая система трубок разного калибра, выполняющая транспортную, трофическую, обменную функцию и функцию регуляции микроциркуляции крови в органах и тканях. Источником развития кровеносных сосудов является мезенхима. На третьей неделе эмбрионального развития вне организма зародыша в стенке желточного мешка и в хорионе (у млекопитающих) образуются скопления клеток мезенхимы – кровяные островки. Периферические клетки островков формируют стенки сосудов, а центрально расположенные мезенхимоциты дифференцируются в первичные клетки крови. Позднее таким же образом сосуды появляются в теле зародыша и устанавливается сообщение между первичными кровеносными сосудами внезародышевых органов и тела зародыша. Дальнейшее развитие сосудистой стенки и приобретение различных особенностей строения происходит под влиянием гемодинамических условий к которым относятся: давление крови, величина его скачков, скорость кровотока. Кровеносные сосуды подразделяются на артерии, вены и сосуды микроциркуляторного русла, к которым относятся артериолы, капилляры, венулы и артериоло-венулярные анастомозы. Общий план строения стенки кровеносных сосудов. За исключением капилляров и некоторых вен, кровеносные сосуды имеют общий план строения, все они состоят из трех оболочек: Внутренняя оболочка (интима) состоит из двух обязательных слоев - эндотелия – непреывного слоя клеток однослойного плоского эпителия, лежащих на базальной мембране и выстилающих внутреннюю поверхность сосуда; - подэндотелиального слоя (субэндотелий), образованного рыхлой волокнистой соединительной тканью. Средняя оболочка в составе которой обычно присутствуют гладкие миоциты и образуемое этими клетками межклеточное вещество, представленное протеогликанами, гликопротеинами, коллагеновые и эластические волокна. Наружная оболочка (адвентиция) представлена рыхлой волокнистой соединительной тканью, с находящимися в ней сосудами сосудов, лимфатическими капиллярами и нервами. 2. Артерии. Классификация. Особенности строения и функции артерий различного типа: мышечного, мышечноэластического и эластического. Артерии – это сосуды, обеспечивающие продвижение крови от сердца к микроциркуляторному руслу в органах и тканях. По артериям течет артериальная кровь, за исключением легочной и пупочной артерий. По количественному соотношению эластических и мышечных элементов в стенке сосуда, артерии подразделяются на: Артерии эластического типа. Артерии смешанного типа (мышечно- эластического) типа. Артерии мышечного типа. Строение артерий эластического типа. К артериям данного типа относятся аорта и легочная артерия. Стенка данных сосудов подвержена большим перепадам давления, поэтому им требуется высокая эластичность. 1. Внутренняя оболочка состоит из трех слоев: - слой эндотелия - подэндотелиальный слой, имеющий значительную толщину, т.к. он амортизирует скачки давления. Представлен рыхлой волокнистой соединительной тканью. В пожилом возрасте здесь появляются холестерин и жирные кислоты. - сплетение эластических волокон, представляет собой густое переплетение продольно и циркулярно расположенных эластических волокон. 2. Средняя оболочка представлена 50-70 окончатыми эластическими мембранами, которые имеют вид цилиндров, вставленных друг в друга, между которыми имеются отдельные гладкие миоциты, эластические и коллагеновые волокна. 3. Наружная оболочка представлена рыхлой волокнистой соединительной тканью с кровеносными сосудами, питающими стенку артерии (сосуды сосудов) и нервами. Строение артерий смешанного (мышечно – эластического) типа. К артериям данного типа относятся подключичная, сонная и подвздошная артерии). 1. Внутренняя оболочка состоит из трех слоев: - эндотелий подэндотелиальный слой - внутренняя эластическая мембрана. 2. Средняя оболочка состоит из примерно равного количества эластических элементов (к которым относятся волокна и эластические мембраны) и гладких миоцитов. 3. Наружная оболочка состоит из рыхлой соединительной ткани, где наряду с сосудами и нервами, находятся продольно расположенные пучки гладких миоцитов. Строение артерий мышечного типа. Это все остальные артерии среднего и малого калибра. 1. Внутренняя оболочка состоит из – эндотелия - подэндотелиального слоя - внутренней эластической мембраны 2. Средняя оболочка имеет наибольшую толщину, представлена в основном спирально расположенными пучками гладких мышечных клеток, между которыми располагаются коллагеновые и эластические волокна. Между средней и наружной оболочками артерии находится слабо выраженная наружная эластическая мембрана. 3.Наружная оболочка представлена рыхлой волокнистой соедини тельной тканью с сосудами и нервами, гладких миоцитов нет. 3. Вены. Классификация. Особенности строения вен различного типа (мышечного и безмышечного). Строение венозных клапанов. Вены – это сосуды, несущие кровь к сердцу. По ним течет венозная кровь, за исключением легочной и пупочной вен. строении ряд особенностей по строению с артериями: Вены имеют больший диаметр. Стенка их более тонкая, легко спадается. Слабо развит эластический компонент и подэндотелиальный слой. Более слабое развитие гладкомышечных элементов в средней оболочке. Хорошо выражена наружная оболочка. Наличие клапанов, которые являются производными внутренней оболочки, снаружи створки клапанов покрыты эндотелием, их толщу образует рыхлая волокнистая соединительная ткань, а в основании находятся гладкие миоциты. Сосуды сосудов содержатся во всех оболочках сосуда. Классификация вен Вены безмышечного типа. Вены мышечного типа, которые в свою очередь подразделяются на: вены со слабым развитием миоцитов - вены со средним развитием миоцитов - вены с сильным развитием миоцитов. Строение вены безмышечного типа. Располагаются вены подобного типа в головном мозге, его оболочках, сетчатке, плаценте, селезенке, костной ткани. Стенка сосуда образована эндотелием, окруженным рыхлой волокнистой соединительной тканью, плотно срастается со стромой органов и поэтому не спадается. Строение вен со слабым развитием миоцитов. Это вены лица, шеи, верхней части тела и верхняя полая вена. 1. Внутренняя оболочка состоит из – эндотелия - слабо развитого подэндотелиального слоя 2. В средней оболочке слабо развитые циркулярно расположенные пучки гладкомышечных клеток, между которыми располагаются значительной толщины прослойки рыхлой соединительной ткани. 3. Наружная оболочка представлена рыхлой волокнистой соединительной тканью. Строение вен со средним развитием миоцитов. К ним относятся плечевая вена и мелкие вены организма. 1. Внутренняя оболочка состоит из: - эндотелия - подэндотелиального слоя 2. Средняя оболочка включает несколько слоев циркулярно расположенных миоцитов. 3. Наружная оболочка толстая, содержит в рыхлой волокнистой соединительной ткани продольно расположенные пучки гладких миоцитов. Строение вен с сильным развитием миоцитов. Располагаются такие вены в нижней части тела и нижних конечностях. Помимо хорошего развития миоцитов во всех слоях стенки характеризуются наличием клапанов, обеспечивающих движение крови в сторону сердца. 4. Сосуды микроциркуляторного русла. Артериолы, венулы, гемокапилляры, их классификация, функции, строение. Микроциркуляторное русло (МЦР) – это система мелких сосудов, которая обеспечивает регуляцию кровенаполнения органов, транскапиллярный обмен и дренажно-депонирующую функцию. Состав МЦР: 1) артериолы, в т.ч. конечные артериолы (диаметр 50-100 мкм), 2) прекапилляры (диаметр 14-16 мкм), 3) гемокапилляры (кровеносные капилляры) (диаметр 3-40 мкм), 4) посткапилляры (диаметр 8-30 мкм), 5) венулы (диаметр от 30 до 100 мкм), 6) артериоловенулярные анастомозы, 7) лимфатические капилляры. Развивается из мезенхимы в стенке желточного мешка и ворсин хориона (вне тела зародыша) на 2-3 неделе эмбрионального развития. Мезенхимные клетки объединяются с образованием кровяных островков. Артериола– мелкий артериальный сосуд мышечного типа, имеющий диаметр 50-100мкм. Стенка состоит из: 1. Внутренняя оболочка: Эндотелия – однослойный, плоский эпителий, ангеодермального типа. Эпителиоциты полигональные, с микровыростами. Субэндотелия – РНСТк. Внутренней эластической мембраны, в которой имеются отверсия, через которые проникают гладкие миоциты и осуществляется работа сосуда. 2. Средняя – 1-2 слоя циркулярно расположенных гладких миоцитов 3. наружняя оболочка образована РНСТк. Функция артериол: транспортная, обменная, обеспечение регуляции кровотока и кровенаполнения органов и тканей. Капилляры имеют различный диаметр в зависимости от функций и располодения в органах. Типы: Соматический – находятся в скелетной МТк легких, кровоснабжают нервную ткань. диаметр 4-7 мкм. Слои: Эндотелий на базальной мембране – непрерывен Клетки-перициты Адвентициальные клетки. Фенестрированные – находтся в составле капиллярного клубочка почки, ворсин кишечника, входят в состав эндокринных органов. Эндотелий истончается в своей цитоплазменной части, т.о. облегчается диффузия. Базальная мембрана непрерывна. Синусоидные – диаметр 2030 мкм. Находится в паренхиме печени, структурах органов кроветворения. Эндотелий прерывистый, прерывная базальная мембрана – содержит поры. Они могут закрываться, т.о. диаметр непостоянный, может выключаться из кровотока. Наличие пор способствует миграции высокомолекулярных соединений и форменных элементов крови. Венулы классифицируются на 3 разновидности: 1) посткапиллярные венулы (диаметр 8-30 мкм); 2) собирательные венулы (диаметр 30-50 мкм); 3) мышечные венулы (диаметр 50-100 мкм). Стенка посткапиллярных венул мало чем отличается от венозного конца капилляра. Разница заключается в том, что в стенке посткапиллярных венул больше перицитов, т. е. в посткапиллярных венулах есть эндотелий и перициты, но нет миоцитов. Стенка собирательных венулотличается появлением в средней оболочке гладких миоцитов и лучше выраженной адвентициальной оболочкой. Стенка мышечных венулхарактеризуется содержанием в средней оболочке 1-2 слоев гладких миоцитов. Функции венул: 1) дренажная (поступление из соединительной ткани в просвет венулы продуктов обмена); 2) из венул в окружающую ткань мигрируют форменные элементы крови. 5. Лимфатические сосуды. Классификация и строение. Лимфатические капилляры, их строение. Участие лимфатических капилляров в системе микроциркуляции. Лимфатические сосуды — часть лимфатической системы, включающей в себя еще и лимфатические узлы. В функциональном отношении лимфатические сосуды тесно связаны с кровеносными, особенно в области расположения сосудов микроциркуляторного русла. Именно здесь происходят образование тканевой жидкости и проникновение ее в лимфатическое русло. Через мелкие лимфоносные пути осуществляются постоянная миграция лимфоцитов из кровотока и их рециркуляция из лимфатических узлов в кровь. Классификация. Среди лимфатических сосудов различают лимфатические капилляры, интра- и экстраорганные лимфатические сосуды, отводящие лимфу от органов, и главные лимфатические стволы тела — грудной проток и правый лимфатический проток, впадающие в крупные вены шеи. По строению различают лимфатические сосуды безмышечного (волокнисто мышечного типов. Лимфатические капилляры. Лимфатические капилляры — начальные отделы лимфатической системы, в которые из тканей поступает тканевая жидкость вместе с продуктами обмена веществ. Лимфатические капилляры представляют собой систему замкнутых с одного конца трубок, анастомозирующих друг с другом и пронизывающих органы. Стенка лимфатических капилляров состоит из эндотелиальных клеток. Базальная мембрана и перициты в лимфатических капиллярах отсутствуют. Эндотелиальная выстилка лимфатического капилляра тесно связана с окружающей соединительной тканью с помощью стропных, или фиксирующих, филаментов, которые вплетаются в коллагеновые волокна, расположенные вдоль лимфатических капилляров. Лимфатические капилляры и начальные отделы отводящих лимфатических сосудов обеспечивают гематолимфатическое равновесие как необходимое условие микроциркуляции в здоровом организме. 6. Сердце. Строение стенки сердца, его оболочек, их тканевой состав. Особенности кровоснабжения и регенерации сердца. Сердечные клапаны. Сердце – основной орган, приводящий в движение кровь. Развитие: первая закладка сердца появляется в начале 3-й недели развития у эмбриона в виде скопления мезенхимных клеток. Позднее эти скопления превращаются в две удлиненные трубочки, впадающие вместе с прилегающими висцеральными листками мезодермы в целомическую полость. Мехенхимные трубочки сливаются – образуется эндокард. Та область висцеральных листков мезодермы, которая прилежит к этим трубочкам, называется миоэпикардиальными пластинками. Из них дифференцируются 2 части – внутренняя, прилежит к мезенхимной трубке – миокард: наружная - эпикард. В стенке сердца различают 3 оболочки: внутреннюю – эндокард, среднюю (мышечную) – миокард , наружную – эпикард. Эндокард напоминает по строению стенку сосуда. В нём выделяют 4 слоя: эндотелийна базальной мембране; подэндотелиальный слой из рыхлой соединительной ткани; мышечно-эластический слой, включающий гладкие миоциты и эластические волокна; наружный соединительнотканный слой .Сосуды имеются лишь в последнем из этих слоёв. Остальные слои питаются путём диффузии веществ непосредственно из крови, проходящей через камеры сердца. В миокарде предсердий различают 2 мышечных слоя: внутренний продольный и наружный циркулярный. В миокарде желудочков - 3 слоя: относительно тонкие внутренний и наружный - продольные, прикрепляющиеся к фиброзным кольцам, окружающим предсердно-желудочковые отверстия; и мощный срединный слой с циркулярной ориентацией. Эпикард включает 3 слоя: а) мезотелий- однослойный плоский эпителий, развивающийся из мезодермы б) тонкую соединительнотканную пластинку, содержащую несколько чередующихся слоёв коллагеновых и эластических волокон и кровеносные сосуды, в) слой жировой ткани. Кровоснабжение сердца осуществляется двумя коронарными артериями - правой и левой. Т.к. они отходят у основания клапана аорты, то при систоле их просвет закрывается створками клапана. От коронарных артерий отходят ветви, питающие отдельные части сердца. Из капилляров кровь собирается в вены, которые впадают не в полые вены, а непосредственно в полость сердца. Клапаны сердца представляют собой складки эндокарда - створки и закрывают предсердно-желудочковые отверстия. Клапан между правым предсердием и правым желудочкам имеет три створки и называется правым предсердножелудочковым(трехстворчатым) клапаном. Левый предсердно-желудочковый клапан двухстворчатый или митральный - это клапан между левым желудочком и левым предсердием. С помощью сухожильных нитей края створок клапанов соединены с сосочковыми мышцами стенок желудочков, что не позволяет своркам выворачиваться в сторону предсердий, и не допускает обратного тока крови из желудочков в предсердие. Около отверстий легочного ствола и аорты так же имеются клапаны в виде трех карманов, открывающихся по направлению тока крови по этим сосудам. Это полулунные клапаны. При уменьшении давления в желудочках сердца они заполняются кровью, их края смыкаются, закрывают просветы легочного ствола и аорты и препятствуют обратному проникновению крови в сердце. 7. Проводящая система сердца, морфофункциональная характеристика. Иннервация. Структурные основы эндокринной функции сердца. Проводящая система сердца — мышечные клетки, формирующие и проводящие импульсы к сократительным клеткам сердца. В состав проводящей системы входят синусно-предсердный (синусный) узел, предсердно-желудочковый узел, предсердно-желудочковый пучок (пучок Гиса) и их разветвления (волокна Пуркинье), передающие импульсы на сократительные мышечные клетки. Клетки узла проводящей системы: формирование импульса происходит в синусном узле (в центре узла – пейсмейкерные клетки, по периферии – переходные клетки – передают возбуждение к клеткам пучка и рабочему миокарду). Клетки пучка Гиса и волокон Пуркинье передают возбуждение переходных клеток клеткам миокарда. Связаны между собой нексусами и десмосомами. ИННЕРВАЦИЯ:в стенке сердца расположены нервные сплетения и ганглии. Рецепторные окончания в стенке сердца (свободные и инкапсулированные) образованы нейронами ганглиев блуждающего нерва и спинномозговых узлов и за счет ветвления дендритов равноотростчатых нейроцитов внутриорганных ганглиев. Функциональные резервы сердца включают его способность функционировать как особый эндокринный орган, имеющий специфические железистые клетки в миокарде предсердий. Они секретируют в кровь полифункциональный гормон –«предсердный натрий-диуретический фактор» (ПНФ), осуществляющий: 1/ регуляцию артериального давления; 2/ расслабление гладких мышц сосудов; 3/ воздействие на почки и регуляцию водного и солевого баланса; 4/ посредничество в регуляции объема циркулирующей крови; 5) воздействие на надпочечники; 6/ регуляцию параметров гемодинамики; XI. ОРГАНЫ ЧУВСТВ 1. Органы чувств. Понятие об анализаторах. Органы чувств — это периферические концы анализаторов. Анализатор — это афферентное звено рефлекторной дуги, включающее чувствительный нейрон органа чувств и ассоциативно-афферентные нейроны, передающие нервный импульс на нейроны коры головного мозга. Анализатор состоит из: 1) периферической (рецепторной) части, в которой расположены рецепторные клетки, являющиеся составной частью органов чувств. Различают механорецепторы (слуха, равновесия, тактильные рецепторы кожи, аппаратадвижения), хеморецепторы (органов вкуса, обоняния, сосудистые интерорецепторы), фоторецепторы, терморецепторы, болевые рецепторы; 2) промежуточной части – цепь вставочных нейронов, по которым нервный импульс от рецепторных клеток передается к корковым центрам; 3) центральной части — коры головного мозга, в которой происходит анализ и синтез полученной информации и готовится ответная реакция. Классификация органов чувств. В зависимости от строения и функции делятся на три типа: 1 тип – первичночувствующие. Рецепторами являются нейросенсорные клетки, т.е. раздражение воспринимают сами нервные клетки. К ним относятся органы зрения и обоняния. 2 тип – вторичночувствующие. Рецепторами являются эпителиальные клетки (сенсоэпителиальные), которые передают раздражение на дендриты чувствительных нейронов. К ним относятся органы равновесия, слуха, вкуса. 3 тип – не имеющие органоспецифического строения. Это орган осязания. 2. Орган вкуса. Общая характеристика. Строение и клеточный состав вкусовых почек. Иннервация. Орган вкуса — периферическая часть вкусового анализатора представлен рецепторными эпителиальными клетками во вкусовых почках. Они воспринимают вкусовые раздражения. Вкусовые почки располагаются в желобоватых, листовидных и фибовидных сосочков языка человека. Развитие. Иннервация вкусовых почек появляется одновременно с возникновением их зачатков. Строение. Каждая вкусовая почка имеет эллипсоидную форму она состоит из 40—60 клеток, 5 видов клеток: сенсоэпителиалъные, «темные» поддерживающие, базальные малодифференцированные и периферические. Сенсоэпителиалъные клетки. Светлые узкие содержат светлое ядро, вокруг которого располагаются митохондрии, органеллы синтеза, первичные и вторичные лизосомы. На цитолемме базальной части клеток берут начало дендриты чувствительных нейронов. Во вкусовых почках передней части языка обнаружен сладкочувствительный рецепторный белок, задней части — горькочувствительный. Поддерживающие эпителиоциты овального ядра с большим количеством гетерохроматина. Около аппарата Гольджи встречаются гранулы, имеются микроворсинки. Базальные малодифференцированные клетки небольшим объемом цитоплазмы вокруг ядра и слабым развитием органелл. В этих клетках выявляются фигуры митоза. Базальные клетки в не достигают поверхности эпителиального слоя. Периферические клетки имеют серповидную форму, содержат мало органелл, но в них много микротрубочек и нервных окончаний. 3. Орган обоняния. Строение и клеточный состав обонятельной выстилки. Основной орган обоняния, являющийся периферической частью сенсорной системы, представлен обонятельной областью. Развитие. Основной орган обоняния имеет эктодермальное происхождение и развивается из плакод. У зародышей человека на 4-м месяце развития из элементов, образуются поддерживающие эпителиоциты и нейросенсорныеобонятельные клетки. Аксоны обонятельных клеток, объединившись между собой, образуют в совокупности 20—40 нервных пучков. Здесь осуществляется синаптический контакт между терминалями аксонов и дендритами митральных нейронов обонятельных луковиц. Строение. Орган обоняния — состоит из пласта многорядного эпителия, в котором различают обонятельные нейросенсорные клетки, поддерживающие и базальные эпителиоциты. Поверхность обонятельной выстилки покрыта слоем слизи. Рецепторные, обонятельные клетки располагаются между поддерживающими эпителиоцитами и отросток — дендрит и длинный — центральный — аксон. Обонятельных клеток, у человека 6 млн. Обонятельными булавами несут до 10—12 обонятельных ресничек. Цитоплазма содержит митохондрии. Периферические отростки обонятельных клеток могут сокращаться под действием пахучих веществ. Ядра обонятельных клеток светлые.В соединительнотканном слое центральные отростки составляют пучки безмиелинового обонятельного нерва, которые объединяются в 20—40 обонятельных нитей и через отверстия решетчатой кости направляются в обонятельные луковицы. Поддерживающие эпителиоциты формируют многорядный эпителиальный пласт, в котором и располагаются обонятельные клетки. В цитоплазме их имеется эндоплазматическая сеть. В цитоплазме поддерживающих клеток содержится коричнево-желтый пигмент. Базальные эпителиоциты находятся на базальной мембране и снабжены цитоплазматическими выростами. Цитоплазма их заполнена рибосомами и не содержит тонофибрилл. 4. Орган зрения. Рецепторный аппарат глаза. Глаз — орган зрения, представляющий собой периферическую часть зрительного анализатора. Развитие. Глаз развивается из различных эмбриональных зачатков. Сетчатка и зрительный нерв формируются из нервной трубки путем образования глазных пузырьков. Часть эктодермы утолщается, давая начало зачатку хрусталика. В процессе развития внутренняя стенка глазного бокала преобразуется в сетчатку, а наружная — в пигментный слой сетчатки. Из нейробластов внутренней стенки глазного бокала образуются колбочковые и палочковые фоторецепторные клетки и другие нейроны сетчатки. Глазное яблоко состоит из трех оболочек. Наружная оболочка, прикрепляются наружные мышцы глаза, обеспечивает защитную функцию. В ней различают передний прозрачный отдел —роговицу и задний непрозрачный отдел — склеру. Средняя оболочка выполняет основную роль в обменных процессах. Она имеет три части: часть радужки, часть цилиарного тела и собственно сосудистую — хориодею. Внутренняя, чувствительная оболочка глаза — сетчатка — сенсорная, рецепторная часть зрительного анализатора, в которой происходят фотохимические превращения зрительных пигментов. Оболочки глаза формируют три функциональных аппарата: светопреломляющий; аккомодационный; рецепторный аппарат. Наружная фиброзная оболочка — склера — образована плотной оформленной волокнистой соединительной тканью, содержащей пучки коллагеновых волокон. Аккомодационный аппарат глаза. Включает радужку, ресничное тело. Функция: 1.Изменение кривизны хрусталика в зависимости от дальности рассматриваемых объектов. 2.Фокусировка изображения на сетчатке. 3.Приспособление к разной интенсивности освещения. Рецепторный аппарат глаза. Представлен зрительной частью сетчатки. В сетчатке различают наружный пигментный слой и внутренний–светочувствительный. В сетчатке выделяют заднюю зрительную часть, цилиарную (покрывает цилиарное тело), радужковую (покрывает заднюю поверхность радужки). В сетчатке различают три типа нейронов. 1.Фоторецепторные нейроны (палочковые и колбочковые). 2.Ассоциативные нейроны сетчатки. 3.Ганглионарные нейроны. 5. Орган зрения. Диоптрический и аккомодационный аппараты глаза. Глаз — орган зрения, представляющий собой периферическую часть зрительного анализатора. Развитие. Глаз развивается из различных эмбриональных зачатков. Сетчатка и зрительный нерв формируются из нервной трубки путем образования глазных пузырьков. Часть эктодермы утолщается, давая начало зачатку хрусталика. В процессе развития внутренняя стенка глазного бокала преобразуется в сетчатку, а наружная — в пигментный слой сетчатки. Из нейробластов внутренней стенки глазного бокала образуются колбочковые и палочковые фоторецепторные клетки и другие нейроны сетчатки. Глазное яблоко состоит из трех оболочек. Наружная оболочка, прикрепляются наружные мышцы глаза, обеспечивает Аккомодационный аппарат глаза. Включает радужку, ресничное тело. Функция: 1.Изменение кривизны хрусталика в зависимости от дальности рассматриваемых объектов. 2.Фокусировка изображения на сетчатке. 3.Приспособление к разной интенсивности освещения. Радужка. Расположена между роговицей и хрусталиком. Строма состоит из рыхлой волокнистой соединительной ткани, много пигментных клеток. Гладкомышечные клетки образуют мышцу, суживающую зрачок и мышцу, расширяющую зрачок, т.е. радужка, является диафрагмой глаза. Слои радужки: 1.Передний эпителий 2.Наружный пограничный (бессосудистый) слой 3.Сосудистый слой. 4.Внутренний пограничный слой 5.Пигментный эпителий. Ресничное тело. Фиксирует хрусталик. Способствует изменению его кривизны. Имеет вид треугольника с основанием, обращенным в переднюю камеру глаза. Ресничная мышца образована пучками гладкомышечных клеток, которые расположены в трех направлениях: продольные волокна, циркулярные волокна, косые волокна. Преобладают волокна, которые при сокращении (видение вблизи) способствуют расслаблению связки и округлению хрусталика. От поверхности цилиарного тела отходят цилиарные отростки. Диоптрический аппарат глаза. Роговица(cornea) - прозрачная оболочка, которая относятся к диоптрическому (светопреломляющему) аппарату глаза. В роговице различают: 1. передний эпителий; 2. переднюю пограничную пластинку (боуменова оболочка); 3. собственное вещество роговицы; 4. заднюю пограничную пластинку (десцеметова оболочка); 5. задний эпителий (эндотелий). Хрусталик (lens) - это прозрачное двояковыпуклое образование, соединенное с цилиарным телом с помощью волокон ресничного пояска. Благодаря этому хрусталик изменяет свою форму при сокращении цилиарной мышцы и, таким образом, является пассивной частью аккомодационного аппарата глаза. Стекловидное тело (corpus vitreum) - это прозрачная масса желеподобного вещества, расположенная в полости между хрусталиком и сетчаткой. На фиксированных препаратах стекловидное тело имеет сетчатое строение. Через стекловидное тело (от диска зрительного нерва до задней поверхности хрусталика) проходит канал- остаток эмбриональной сосудистой системы глаза. 6. Орган слуха. Наружное ухо, среднее ухо, внутреннее ухо (костный и перепончатый лабиринты). Улитковая часть перепончатого лабиринта. Гистофизиология восприятия звуков. Органом равновесия и слуха служит ухо. Среднее ухо состоит из барабанной полости, слуховых косточек и слуховой трубы. Барабанная полость — покрытое однослойным плоским эпителием. В ней имеются два отверстия (окна). В овальном окне располагается основание стремечка. Овальное окно отделяет барабанную полость от вестибулярной лестницы улитки. Второе окно круглое, закрыто волокнистой мембраной. Круглое окно отделяет барабанную полость от барабанной лестницы улитки. Слуховые косточки — молоточек, наковальня, стремечко как система рычагов передают колебания барабанной перепонки наружного уха к овальному окну, от которого начинается вестибулярная лестница внутреннего уха. Слуховая труба, соединяющая барабанную полость с носовой частью глотки, имеющие бокаловидные железистые клетки. Через слуховую трубу регулируется давление воздуха в барабанной полости среднего уха. Внутреннее ухо состоит из костного лабиринта и расположенного в нем перепончатого лабиринта, в котором находятся рецепторные клетки — волосковые сенсорные эпителиоциты органа слуха и равновесия. Слуховые рецепторные клетки — в спиральном органе улитки, а рецепторные клетки органа равновесия — в эллиптическом и сферическом мешочках и ампулярных гребешках полукружных каналов. Улитковый канал представляет собой спиральный слепо заканчивающийся мешок, заполненный эндолимфой и окруженный снаружи перилимфой. Улитковый канал на поперечном разрезе имеет форму треугольника, стороны которого образованы вестибулярной мембраной, сосудистой полоской, лежащей на наружной стенке костной улитки, и базилярной пластинкой. Вестибулярная мембрана образует верхнемедиальную стенку канала. Наружная стенка образована сосудистой полоской. Наружное ухо включает (1) ушную раковину, (2) наружный слуховой проход и (3) барабанную перепонку. 1. Ушная раковина состоит из эластического хряща, покрытого кожей с волосами, сальными железами и небольшим числом эккринных потовых, желез. 2. Наружный слуховой проход выстлан кожей, содержащей щетинистые волосы, сальные и церуминозные (видоизмененные апокрин-ные потовые) железы, выделяющие ушную серу. 3. Барабанная перепонка отделяет наружное ухо от среднего и передает воздушные звуковые колебания на слуховые косточки. Представляет собой пластинку из соединительной ткани, которая содержит фибробласты, коллагеновые и эластические волокна, образующие наружный радиальный и внутренний циркулярный слои. Гистофизиология органа слуха. Колебания воздуха передаются на барабанную перепонку и через цепочку слуховых косточек достигают основы стремени. Смещаясь подобно поршню в овальном окне основа стремени передает колебания на перилимфу вестибулярных лестницы улитки. Через отверстие на вершине улитки колебания переходят на перилимфу барабанной лестницы. Отдача колебаний происходит через круглое окно, которое выпячивается в барабанную полость в то время, когда основа стремени углубляется в овальное окно. Колебания перилимфы вестибулярной лестницы передаются через вестибулярную мембрану на эндолимфу протока завитки и охватывают базилярную и текториальну мембраны. Эти движения соответствуют частоте и интенсивности звуков. В результате происходит отклонение стереоцилий сенсорных клеток и их возбуждение. Оно сопровождается взаимодействием ацетилхолина, содержащегося в эндолимфе, с холинорецепторным белком в мембранах стереоцилий. Это приводит к возникновению рецепторного потенциала (микрофонный эффект). Нервные импульсы через слуховой нерв передаются в центральные отделы слухового анализатора 7. Орган равновесия. Вестибулярная часть перепончатого лабиринта: эллиптический и сферический мешочки и полукружные каналы. Их рецепторные отделы: строение и клеточный состав пятна и ампулярных гребешков. Гистофизиология вестибулярного лабиринта. Орган равновесия включат специализированные рецепторные зоны в мешочке, маточке и ампулах полукружных каналов. Мешочек и маточка содержат пятна(макула)-участки в которых однослойный плоский эпителий перепончатого лабиринта сменяется призматически. Макулы включпет 7.5-9 тыс. сенсорноэпителиальных клеток, связанных комплексами соединений с поддерживающими клетками и покрытых отолитовой мембраной. Макула маточки занимает горизонтальное положение, а макула мешочка-вертикальное. Сенсорно-эпителиальные клетки содержат многочисленные митохондрии, развитую аЭПС и крупный комплекс Гольджи, на апикальной полюсе распологаются одна эксцентрично лежащая ресничка и 40-80 жестких стереоцилий различной длины. Ампулы полукружных каналов образуют выступы-ампулярные гребешки, располагающиеся в плоскости перпендикулярной оси канала. Гребешки выстланы призматическим эпителием, содержащим клетки тех же типов что и макулы. Ампулярные гребешки воспринимают угловые ускорения: при вращение тела возникает ток эндолимфы, который отклоняет купол, что стимулирует волосковые клетки вследствие изгибаний стереоцилий. ВЕСТИБУЛЯРНАЯ ЧАСТЬ ПЕРЕПОНЧАТОГО ЛАБИРИНТА - имеет 2 расширения: 1. Мешочек - сферической формы расширение. 2. Маточка - расширение эллептической формы. Эти два расширения соединены друг с другом тонким канальцем. С маточкой связаны три взаимоперпендикулярные полукружные каналы с расширениями - ампулами. Большая часть внутренней поверхности мешочка, маточки и полукружных каналов с ампулами покрыта однослойным плоским эпителием. В тоже время в мешочке, маточке и в ампулах полукружных каналов имеются участки с утолщенным эпителием. Эти участки с утолщенным эпителием в мешочке и маточке называются пятнами или макулами, а в ампулах - гребешками или кристами. XII. ОРГАНЫ КРОВЕТВОРЕНИЯ И ИММУННОПОЭЗА 1. Морфофункциональная характеристика первичных и вторичных органов иммуногенеза. К системе органов кроветворения и иммунной защиты относят красный костный мозг, тимус (вилочковая железа), селезенку, лимфатические узлы, а также лимфатические узелки пищеварительного тракта (миндалины, лимфатические узелки кишечника) и других органов. Различают центральные (первичные) и периферические (вторичные) органы кроветворения и иммунной защиты. К центральным органам кроветворения у человека относятся красный костный мозг и тимус. В красном костном мозге образуются эритроциты, кровяные пластинки (тромбоциты), гранулоциты и предшественники лимфоцитов. Тимус — центральный орган лимфопоэза. В периферических кроветворных органах (селезенка, лимфатические узлы, гемолимфатические узлы) происходят размножение приносимых сюда из центральных органов Т- и В-лимфоцитов и специализация их под влиянием антигенов в эффекторные клетки, осуществляющие иммунную защиту, и клетки памяти (КП). Кроме того, здесь погибают клетки крови, завершившие свой жизненный цикл. Органы кроветворения функционируют содружественно и обеспечивают поддержание морфологического состава крови и иммунного гомеостаза в организме. Координация и регуляция деятельности всех органов кроветворения осуществляются посредством гуморальных и нервных факторов организма, а также внутриорганных влияний, обусловленных микроокружением. Несмотря на различия в специализации органов гемопоэза, все они имеют сходные структурно-функциональные признаки. В основе большинства их лежит ретикулярная соединительная ткань, которая образует строму органов и выполняет роль специфического микроокружения для развивающихся гемопоэтических клеток и лимфоцитов. В этих органах происходят размножение кроветворных клеток, временное депонирование крови или лимфы. Кроветворные органы благодаря наличию в них специальных фагоцитирующих и иммунокомпетентных клеток осуществляют также защитную функцию и способны очищать кровь или лимфу от инородных частиц, бактерий и остатков погибших клеток. 2. Строение костного мозга, функции. Красный костный мозг — это центральный орган кроветворения, в котором из СКК(стволовых кроветворных клеток) развиваются эритроциты, нейтрофильные, эозинофильные и базофильные гранулоциты, моноциты, В-лимфоциты, предшественники Т-лимфоцитов и тромбоциты. В красном костном мозге происходит антигеннезависимая дифференцировка В-лимфоцитов. Клетки микроокружения красного костного мозга представлены ретикулоцитами, макрофагами, остеогенными клетками и адипоцитами. Все клетки микроокружения редко делятся. Развитие. ККМ закладывается в конце 1 месяца из мезенхимы. 1е клетки появляются в ключице эмбриона (2 мес), далее в плоских костях (3 мес), трубчатых (4 мес). ККМ уходит в эпифизы, а диафизы заполняются ЖКМ.(желт.кост.мозг). На 5-6-м месяце окончательно формируется костномозговая полость (с помощью остеокластов) в диафизах трубчатых костей, и с этого момента красный костный мозг становится основным органом кроветворения. Строение. ККМ состоит из компонентов: 1.Стромальный (ретикулярная ткань, ретикулярные волокна, которые соединяются с костными трабекулами, а с другой стороны подходят к кровеносным сосудам и образуют сеть, в стенке которых содержится гемопоэтический компонент – островок кроветворения) 2.Сосудистый (капилляры распадаются на посткапиллярые синусы в костно-мозговой полости снабжены сфинктерами – происходит выключение синусов из кровотока) 3.Гемопоэтический (миелопоэз, лимфопоэз) Функция: Главной функцией красного костного мозга является образование клеток крови, разрушение изношенных красных кровяных клеток и накопление (в макрофагах) железа, образовавшегося при распаде гемоглобина. Регенерация. После удаления части красного костного мозга его ретикулярная строма восстанавливается за счет пролиферации оставшихся недифференцированных ретикулярных клеток, а гемопоэтические клетки — за счет вселения стволовых клеток. 3. Тимус. Строение и тканевой состав коркового и мозгового вещества долек. Васкуляризация. Строение и значение гематотимического барьера. Инволюция тимуса. Тимус (вилочковая железа) Это центральный орган кроветворения и иммунитета. В эмбриогенезе образуется из эпителия глоточной кишки (3-я, 4-я пара жаберных карманов). Эпителий разрастается и постепенно разделяется на дольки, между которыми из мезенхимы образуются соединительнотканные перегородки. Т.о., стромой каждой дольки является эпителий, который, потеряв строение пласта, постепенно разрыхляется и принимает ретикулоподобный вид, поэтому клетки называются ретикулоэпителиоцитами. В дольке могут располагаться макрофаги, сюда врастают сосуды со своим эндотелием и адвентициальными клетками, что составляет микроокружение для созревания популяций лимфоцитов. На уровне полустволовых клетки заселяют дольки тимуса. Здесь происходит их дифференцировка и образование на Т-лимфоцитах специальных рецепторов. Строение тимуса Тимус окружен снаружи соединительнотканной капсулой. Анатомически подразделяется на левую и правую доли и перешеек. Прослойки соединительной ткани разделяют его на дольки. Каждая долька подразделяется на две зоны: 1. Корковое вещество: эпителиальные клетки располагаются более рыхло и соединяются между собой с помощью длинных отростков. 2. Мозговое вещество: эпителиальные клетки лежат более компактно. Лимфоциты в первую очередь заселяют мозговое вещество. Но затем в эмбриогенезе они сосредоточены на 95% в корковом веществе (имеет более темный цвет). В корковом веществе по периферии располагаются бластные клетки (лимфобласты) – это т.н. субкапсулярная зона. Здесь сосредоточено 5% Т-лимфоцитов, которые устойчивы (резистентны) к физическим факторам, облучениям и глюкокортикоидам коры надпочечников. При облучении или при стрессе лимфоциты данной зоны меньше всего страдают, в то время как остальные лимфоциты могут разрушаться (при стрессе). Кровоснабжение тимуса: Корковое и мозговое вещества кровоснабжаются отдельно. При этом Т-лимфоциты из коркового вещества не проходят в мозговое, они могут мигрировать в Т-зоны периферических органов кроветворения. Кровоснабжение мозгового вещества более замкнуто, поэтому из него не могут выйти Т-лимфоциты. Этому препятствует специальный барьер, который представлен эндотелиальными клетками и базальной мембраной капилляров, эпителиальными клетками стромы и макрофагами, имеющимися здесь. Регенерация. Возможна только в детском возрасте. Мокроокружением тимуса вырабатываются факторы способствующие кроветворению – тимозины: Т-активин и В-активин. 4. Лимфатические узлы. Морфофункциональная характеристика. Корковое и мозговое вещество. Т- и Взависимые зоны. Система синусов. ЛИМФАТИЧЕСКИЕ УЗЛЫ – более крупные органы от 2 мм до 1,5 см и более. Они располагаются в анатомически определенных местах. Развиваются из мезенхимы на 2 мес внутриутробной жизни. В эмбриогенезе идет универсальное кроветворение. С формированием капсул и трабекул из волокнистой соединительной ткани меняется микроокружение. Изменяется тип на антигензависимый лимфоцитопоэз. Имеют бобовидную форму, снаружи – капсула, которая сращена снаружи с окружающей тканью. Внутрь от капсулы отходят трабекулы, в которых проходят кровеносные сосуды. В капсулах и трабекулах имеются гладко-мышечные клетки (выталкивают депонированную лимфу). В строму узла входит ретикулярная ткань, среди ретикулярных клеток есть (макрофаг лимфоузла) – резидентные макрофаги. Встречаются нефагоцитирующие макрофаги: - дендритные; - интердигитирующие. Сами они не фагоцитируют, но способны накапливать на поверхности антиген, при определенном количестве которого они приводят в состояние пролиферации и бласттрансформации лимфоциты (дендритные – В, интердигитирующие – Т). Лимфоидная ткань в лимфатическом узле расположена неравномерно. По периферии располагаются лимфатические узелки. А в мозговом веществе образуют мякотные тяжи. Лимфатические узелки и мякотные тяжи составляют В-зависимые зоны. На границе коркового и мозгового вещества выделяют паракортикальную зону – Т-зависимая зона (Т-лимфоциты). Эти зоны зависят от особого микроокружения, кроме ретикулярных клеток входят нефагоцитирующие макрофаги. Лимфа притекает в лимфатический узел по сосудам с наружной выпуклой стороны узла. Затем попадает в краевой (подкапсулярный) синус. Затем лимфа растекается по щелям, где формирует вокругузелковые синусы, лимфа растекается и по щелям, которые формируют промежуточные (мозговые) синусы. В конце концов лимфа попадает в область ворот, где локализуется воротный синус, откуда идет отток лимфы. Все эти синусы, не являясь сосудами, имеют ретикулярную выстилку. Ретикулиновые волокна перегораживают ход лимфы. Т.о., лимфатический узел выполняет: 1) защитную (барьерную) функцию за счет ретикулярной ткани и макрофагов; 2) кроветворную (только лимфоцитопоэз); 3) иммунобиологическую (в мякотных тяжах В-лимфоциты, сильно активируясь антигеном и Тлимфоцитами превращается на 80% в плазматические клетки, которые вырабатывают антитела); 4) депонирующую лимфу функцию. Регенерация высокая, но при условии сохранения приносящих и выносящих лимфатических сосудов. 5. Селезенка. Строение и тканевой состав (белая и красная пульпа. Т- и Взависимые зоны). Кровоснабжение селезенки. Структурные и функциональные особенности венозных синусов. СЕЛЕЗЕНКА – крупный орган периферической системы кроветворения, образуется из мезенхимы на 2 мес. внутриутробного развития. Строма – ретикулярная ткань. В эмбриогенезе – универсальный орган кроветворения. К концу эмбриогенеза формируется мощная соединительнотканная капсула, от которой внутрь отходят трабекулы. Вследствие меняется микроокружение, и кроветворение затухает, переходя в красный костный мозг. Снаружи капсулы располагается висцеральный листок брюшины. В капсуле и в трабекулах располагаются в большом количестве гладкомышечные клетки, сокращение которых приводит к выбросу депонированной крови. В строму входят ретикулярные клетки, селезеночные макрофаги. Все вещество селезенки подразделяется на: 1) белая пульпа (лимфатические узелки разбросаны по всей селезенке равномерно) 2) красная пульпа (все остальное). Белая пульпа построена более сложно, чем в лимфатическом узле. В ней выделяют: центральную артерию (расположена сбоку), вокруг которой выделяют периартериальную зону (тимус-зависимую). Здесь микроокружение – Т-лимфоциты, строма – ретикулярные клетки и интердигитирующие макрофаги. Центральную часть занимают молодые клетки – Влимфоциты. В В-зоне микроокружение представлено ретикулярными клетками и дендритными макрофагами. Под действием накопленного антигена формируются реактивные центры (светлые), т.е. вокруг светлого центра расположена мантийная зона В-лимфоцитов (и, может быть, Т-лимфоцитов на пути миграции). Снаружи в лимфатическом узелке располагается краевая (маргинальная) зона (В- и Т-лимфоциты на путях миграции). Красная пульпа представлена резидентными макрофагами, хорошо развита синусоидная сосудистая сеть. Селезенка является кроветворным органом. Она выполняет функции: 1) барьерную; 2) иммуно-биологическую; 3) вырабатывает различные поэтины: - тромбоцитопоэтины блокирующего ряда, - эриропоэтины; Селезенка хорошо регенерирует – быстро формируется масса селезенки, но необходимо сохранить сосудистую систему в области ворот. При разрыве селезенки еѐ удаляют из-за больших кровопотерь. 6. Понятие о единой иммунной системе слизистых оболочек: лимфатические узелки в стенках воздухоносных путей, пищеварительного тракта и других органов. Иммунная система - это совокупностm всех лимфоидных органов и скоплений лимфатических клеток тела. Синонимом иммунной системы является лимфатическая система. Лимфоидные органы - это функциональные тканевые образования, в которых образуются иммунные клетки и где они приобретают иммунную специфичность. Среди органов иммунной системы различают: 1. Центральые: вилочковая железа (тимус), костный мозг, бурса (у птиц). 2. Периферические: кровь, лимфа, селезенка, лимфатические узлы. 3. Система лимфоэпителиальных образований: скопления лимфоидной ткани слизистых оболочек желудочно-кишечного тракта, дыхательных и мочеполовых путей. Иммунная система слизистых оболочек формирует защитный барьер, предохраняющий организм человека от болезнетворного воздействия патогенной и условно-патогенной микрофлоры. Защитные реакции, развиваемые иммунной системой слизистых, в норме протекают на фоне минимальной воспалительной реакции и не сопровождаются повреждением окружающих тканей. Это достигается, главным образом, благодаря преимущественной секреции иммунокомпетентными клетками IgA, который способен нейтрализовать патоген и разрушать иммунные комплексы без привлечения комплемента. Следует отметить, что иммунокомпетентные клетки слизистых тесно взаимодействуют с эпителиоцитами, нервными, мышечными и стромальными клетками. Нарушения в системе этих взаимодействий ведут, как правило, к развитию иммуновоспалительных реакций или атрофическим процессам в слизистой. В свою очередь, развитие хронической воспалительной реакции способно приводить к нарушению функционирования иммунной системы слизистых и нарушению местного гомеостаза. Иммунная система слизистых представлена регионарными лимфатическими узлами (в кишечнике – это брыжеечные лимфатические узлы), миндалинами глоточного лимфоидного кольца, лимфоидными (пейеровыми) бляшками кишечника, многочисленными лимфоидными узелками, расположенными в слизистых оболочках, диффузной лимфоидной тканью, а также лимфоидными клетками собственной пластинки (Lamina propria) и межэпителиальными лимфоцитами. XIII. ЭНДОКРИННАЯ СИСТЕМА 1. Нейросекреторные отделы гипоталамуса. Нейроэндокринные нейроны крупноклеточных и мелкоклеточных ядер гипоталамуса. Связь гипоталамуса с адено- и нейрогипофизом. Гипоталамоаденогипофизарная и гипоталамонейрогипофизарная системы. Эндокринная система — совокупность структур: органов, частей органов, отдельных клеток, секретирующих в кровь и лимфу гормоны. В состав эндокринной системы входят специализированные эндокринные железы, или железы внутренней секреции, лишенные выводных протоков, но обильно снабженные сосудами микроциркуляторного русла, в которые выделяются продукты секреции этих желез. Различают центральные и периферические отделы: I. Центральные регуляторные образования эндокринной системы 1. Гипоталамус (нейросекреторные ядра) 2. Гипофиз (аденогипофиз и нейрогипофиз) 3. Эпифиз II. Периферические эндокринные железы 1. Щитовидная железа 2. Околощитовидные железы 3. Надпочечники (корковое и мозговое вещество) Гипоталамус является высшим нервным центром регуляции эндокринных функций. Он контролирует висцеральные функции организма и объединяет эндокринные механизмы регуляции с нервными, будучи мозговым центром симпатического и парасимпатического отделов вегетативной нервной системы. Субстратом объединения нервной и эндокринной систем являются нейросекреторные клетки, которые располагаются в нейросекреторных ядрах гипоталамуса. В гипоталамо-аденогипофизарной системе аккумулируются аденогипофизотропные нейрогормоны — либерины и статины, которые затем поступают в портальную систему гипофиза. В гипоталамо-нейрогипофизарной системе нейрогемальным органом оказывается нейрогипофиз (задняя доля гипофиза), где аккумулируются вазопрессин и окситоцин), выделяемые в кровь. Секреторные нейроны расположены в ядрах серого вещества гипоталамуса. В переднем гипоталамусе располагаются парные супраоптические и паравентрикулярные ядра, которые образованы крупными холинергическими нейросекреторными клетками. В ядрах среднего гипоталамуса адренергические нейросекреторные клетки вырабатывают аденогипофизотропные нейрогормоны, с помощью которых гипоталамус контролирует гормонообразовательную деятельность аденогипофиза. Эти нейрогормоны разделяются на либерины, стимулирующие выделение, продукцию гормонов передней и средней долей гипофиза и статины, угнетающие функции аденогипофиза. Гипофиз состоит из аденогипофиза (передняя доля) и нейрогипофиза (задняя доля). Нейрогипофиз: гормоны не синтезируются: здесь лишь происходит поступление в кровь нейрогормонов, образованных в гипоталамусе, АДГ и окситоцина. Аденогипофиз: Гормоны:Гонадотропные гормоны (стимулируют гонады): фолликулостимулирующий гормон (ФСГ), лютеинизирующий гормон (ЛГ), или лютропин, лактотропный гормон (ЛТГ), пролактин, или лютеотропный гормон.\ 2. Эпифиз (шишковидная железа). Строение, клеточный состав, функции. Эпифиз (шишковидное тело) Развитие. У зародыша человека эпифиз закладывается на 5-6-й неделе внутриутробного периода в виде выпячивания промежуточного мозга. Строение. Снаружи эпифиз окружен тонкой соединительнотканной капсулой, от которой внутрь железы отходят разветвляющиеся перегородки, образующие ее строму и разделяющие паренхиму на дольки. В паренхиме органа различаются клетки двух типов: А) секретообразующие пинеалоциты располагаются в центральной части долек. Это относительно крупные клетки с отходящими от их тела длинными отростками, которые булавовидно расширяясь, направляются к капиллярам и контактируют с ними. Среди пинеалоцитов различают: светлые пинеалоциты, характеризующиеся светлой гомогенной цитоплазмой; темные пинеалоциты, меньшего размера с ацидофильными (а иногда базофильными) включениями в цитоплазме. Обе названные формы не являются самостоятельными разновидностями, а представляют собой клетки, находящиеся в различных функциональных состояниях, или подвергающиеся возрастным изменениям. Б) глиальные клетки преобладают на периферии долек. Их цитоплазма скудна, ядра уплотнены, а отростки направляются к междольковым соединительнотканным перегородкам, образуя своего рода краевую кайму дольки. Функции эпифиза: 1) участвует в регуляции процессов, протекающих в организме ритмически, циклически (например овариально-менструального цикла) или закономерно изменяющихся на протяжении суток (смена дня и ночи); 2) замедляет развитие половой системы; 3) вырабатывает серотонин, который превращается в мелатонин. Этот нейроамин, по-видимому, ослабляет или угнетает секрецию гонадолиберина гипоталамусом и гонадотропинов передней доли гипофиза; 4) продуцируeт ряд белковых гормонов и в их числе: антигонадотропин, ослабляющий секрецию лютропина передней доли гипофиза. гормон, повышающий уровень калия в крови; 5) секретирует около 40 регуляторных пептидов (тироли6ерин, люлиберин и др.) Иннервация. Симпатические нервные волокна. 3. Гипофиз. Строение, функции аденогипофиза, нейрогипофиза, средней доли гипофиза. Связь гипофиза с гипоталамусом. Васкуляризация и иннервация гипофиза. В переднем гипоталамусе располагаются парные супраоптические и паравентрикулярные ядра, которые образованы крупными холинергическими нейросекреторными клетками. В ядрах среднего гипоталамуса адренергические нейросекреторные клетки вырабатывают аденогипофизотропные нейрогормоны, с помощью которых гипоталамус контролирует гормонообразовательную деятельность аденогипофиза. Эти нейрогормоны разделяются на либерины, стимулирующие выделение, продукцию гормонов передней и средней долей гипофиза и статины, угнетающие функции аденогипофиза. Гипофиз состоит из аденогипофиза (передняя доля) и нейрогипофиза (задняя доля). Нейрогипофиз: гормоны не синтезируются: здесь лишь происходит поступление в кровь нейрогормонов, образованных в гипоталамусе, АДГ и окситоцина. Три компонента. В задней доле гипофиза нет секреторных клеток. Имеются три компонента. Питуициты - мелкие глиальные клетки, имеют многочисленные отростки, образующие строму. Кровеносные сосуды - многочисленны, среди них преобладают капилляры. Аксоны нервных клеток гипоталамуса - образуют многочисленные пучки и кончаются накопительными тельцами. Аденогипофиз: Гормоны:Гонадотропные гормоны (стимулируют гонады): фолликулостимулирующий гормон (ФСГ), лютеинизирующий гормон (ЛГ), или лютропин, лактотропный гормон (ЛТГ), пролактин, или лютеотропный гормон. Действие: ФСГ стимулирует в яичниках - рост фолликулов, в семенниках - рост семенных канальцев и сперматогенез. ЛГ стимулирует в яичниках - окончательное созревание фолликула и секрецию эстрогенов, в семенниках - секрецию тестостерона. ЛТГ стимулирует выработку прогестерона жёлтым телом яичника, секреторную активность молочных желёз. Гормоны: Гормоны, стимулирующие другие (не половые) железы: тиреотропный гормон (ТТГ), адренокортикотропный гормон (АКТГ). Действие: ТТГ стимулирует образование и секрецию гормонов щитовидной железы (тироксина и др.). АКТГ стимулирует образование гормонов в коре надпочечников. 4. Щитовидная железа. Строение, функции. Цитофункциональная характеристика фолликулярных и парафолликулярных эндокриноцитов. Щитовидная железа располагается в области шеи кпереди от гортани и состоит из двух долей, соединенных перешейком. В раннем эмбриональном периоде она развивается из энтодермы начальной части первичной кишки. Ее функция заключается в синтезе гормонов тироксина (Т ) и трийодтиронина (Т3), которые необходимы для роста и дифференцировки клеток, регуляции потребления кислорода и основного обмена в организме. Гормоны щитовидной железы влияют на обмен белков, липидов и углеводов. Ткань щитовидной железы состоит из 20—30 млн микроскопических сферических структур, известных как фолликулы щитовидной железы, или тироидные фолликулы. Фолликулы выстланы однослойным эпителием, а находящаяся внутри полость содержит желеобразное вещество — коллоид. Щитовидная железа является единственной эндокринной железой, секреторный продукт которой запасается в очень больших количествах. Этот процесс необычен еще и тем, что гормоны накапливаются в коллоиде внеклеточно. Щитовидная железа – содержит 2 типа эндокринных клеток: фолликулярные эндокриноциты, тироциты (вырабатывают тироксин), и парафолликулярные эндокриноциты (вырабатывают кальцитонин). Фолликулярные эндокриноциты (тироциты) – железистые клетки, составляют большую часть стенки фолликулов. Белковые продукты, синтезируемые тироцитами, выделяются в полость фолликула, где завершается образование йодированных тирозинов и тиронинов (аминокислот). Парафолликулярные эндокриноциты (кальцитониноциты) – в стенке фолликулов, в межфолликулярных прослойках соединительной ткани. Клетки не поглощают йод, но совмещают образование нейроаминов (норадреналин и серотонин) с синтезом белковых гормонов (кальцитонина и соматостатина). Основной функцией щитовидной железы является выработка гормонов: трийодтиронина (обычно обозначается как Т3) и тетрайодтиронина (он же тироксин - Т4). Трийодтиронин является более активным гормоном, в то время как тироксин служит в организме своеобразным "запасом". При необходимости, от Т4 отщепляется одна молекула йода, и он превращается в активный гормон Т3. 5. Околощитовидные железы. Участие околощитовидной железы в регуляции кальциевого гомеостаза. Околощитовидные железы: Функциональное значение околощитовидных желез заключается в регуляции метаболизма кальция. Они вырабатывают белковый гормон паратирин, который стимулирует резорбцию кости остеокластами, повышая уровень кальция в крови, и снижает уровень фосфора в крови, тормозя его резорбцию в почках, уменьшает экскрецию кальция почками. Железа окружена соединительнотканной капсулой. Паренхима представлена трабекулами — скоплениями эпителиальных клеток — паратироцитов. Различают главные паратироциты и оксифильные паратироциты. Главные клетки секретируют паратирин. На секреторную активность околощитовидных желез не оказывают влияния гипофизарные гормоны. Околощитовидная железа по принципу обратной связи быстро реагирует на малейшие колебания в уровне кальция в крови. Ее деятельность усиливается при гипокальциемии и ослабляется при гиперкальциемии. Паратироциты обладают рецепторами, способными непосредственно воспринимать прямые влияния ионов кальция на них. 6. Надпочечники. Морфофункциональная характеристика коркового и мозгового вещества надпочечников. Надпочечники — это эндокринные железы, которые состоят из двух частей - коркового и мозгового вещества, обладающих различным происхождением, структурой и функцией. Снаружи надпочечники покрыты соединительнотканной капсулой, в которой различаются два слоя - наружный (плотный) и внутренний (более рыхлый). От капсулы в корковое вещество отходят тонкие трабекулы, несущие сосуды и нервы. Корковое вещество надпочечников занимает большую часть железы и выделяет кортикостероиды - группу гормонов, влияющих на различные виды обмена, иммунную систему, течение воспалительных процессов. Функция коры надпочечников контролируется адренокортикотропным гормоном гипофиза (АКТГ), а также гормонами почек - ренинангиотензиновой системой. В мозговом веществе продуцируются катехоламины (адреналин, или эпинефрин, и норадреналин, или норэпинефрин), которые влияют на быстроту сердечных сокращений, сокращение гладких мышц и метаболизм углеводов и липидов. 7. Диффузная эндокринная система, локализация элементов, их клеточный состав. Роль гормонов в местной и общей регуляции (на конкретном примере). Эндокринная система — совокупность структур: органов, частей органов, отдельных клеток, секретирующих в кровь и лимфу гормоны. Гормоны — это высокоактивные регуляторные факторы, оказывающие стимулирующее или угнетающее влияние преимущественно на основные функции организма: обмен веществ, соматический рост, репродуктивные функции. Эндокринная система совместно с нервной системой осуществляет регуляцию и координацию функций организма. В состав эндокринной системы входят специализированные эндокринные железы, или железы внутренней секреции, лишенные выводных протоков, но обильно снабженные сосудами микроциркуляторного русла, в которые выделяются продукты секреции этих желез. Одиночные эндокринные клетки рассеяны по разным органам и тканям организма. Подавляющее большинство гормонов принадлежит к белкам (пептиды, олигопептиды, гликопептиды) и производным аминокислот, часть — к стероидам (половые гормоны и гормоны коры надпочечников). Различают центральные и периферические отделы: I. Центральные регуляторные образования эндокринной системы 1. Гипоталамус (нейросекреторные ядра) 2. Гипофиз (аденогипофиз и нейрогипофиз) 3. Эпифиз II. Периферические эндокринные железы 1. Щитовидная железа 2. Околощитовидные железы 3. Надпочечники (корковое и мозговое вещество) III. Органы, объединяющие эндокринные и неэндокринные функции 1. Гонады (семенники, яичники) 2. Плацента 3. Поджелудочная железа IV.Одиночные гормонпродуцирующие клетки –эндокринные клетки в разных отделах пищеварительной, дыхательной и нервной систем. 1. Нейроэндокринные клетки группы неэндокринных органов — APUD-серия 2. Одиночные эндокринные клетки, продуцирующие стероидные и другие гормоны. Совокупность одиночных гормонпродуцирующих клеток называют диффузной эндокринной системой (ДЭС). Среди одиночных гормонпродуцирующих клеток различают две самостоятельные группы: I — нейроэндокринные клетки APUD-серии (нервного происхождения); II — клетки не нервного происхождения. XIV. ПИЩЕВАРИТЕЛЬНАЯ СИСТЕМА 1. Общий план строения стенки пищеварительного тракта. Типы слизистых оболочек. Пищеварительная система человека представляет собой пищеварительную трубку с расположенными рядом с ней (железами (слюнными железами, печенью и поджелудочной железой), секрет которых участвует в процессе пищеварения. Процессом пищеварением называются процессы химической и механической обработки пищи с последующим всасыванием продуктов ее расщепления. Роль желудочно-кишечного тракта в организме человека очень велика: из него идет поступление веществ, обеспечивающих организм необходимой энергией и строительными материалами для восстановления постоянно разрушающихся его структур. Слизистая оболочка. Состоит из эпителия, собственной пластинки слизистой оболочки и мышечной пластинки слизистой оболочки. Эпителий в переднем и заднем отделах пищеварительной трубки многослойный плоский, а в среднем отделе, т. е. в желудке и кишечнике, — однослойный цилиндрический. Железы расположены либо эндоэпителиально, либо экзоэпителиально, либо за пределами пищеварительного канала. В состав слизистой оболочки входит собственная пластинка, которая лежит под эпителием, отделена от него базальной мембраной и представлена рыхлой волокнистой неоформленной соединительной тканью. В ней проходят кровеносные и лимфатические сосуды, нервные элементы, скопления лимфоидной ткани. Расположением мышечной пластинки слизистой оболочки является граница с подслизистой основой. Данная пластинка состоит из нескольких слоев, образованных гладкими мышечными клетками. Подслизистая основа представлена рыхлой волокнистой неоформленной соединительной тканью, она как бы соединяет слизистую оболочку с подлежащими образованиями (мышечной оболочкой или костной основой). Благодаря ей слизистая оболочка имеет подвижность и может образовывать складки. Мышечную оболочку составляет гладкомышечная ткань, в этом случае расположение мышечных волокон может быть циркулярным (внутренний слой) и продольным (наружный слой). Брюшина состоит из соединительнотканной основы, в которой имеются сосуды и нервные элементы, и из мезотелия, окружающего ее снаружи. Кровоснабжение. В тонкой кишке артериальные сплетения формируются также в мышечной оболочке. Капиллярные сети образуются под эпителием слизистой оболочки, вокруг желез, крипт, желудочных ямок, внутри ворсинок, сосочков языка и в мышечных слоях. Вены также формируют сплетения подслизистой основы и слизистой оболочки. 2. Ротовая полость: губы, десны, язык. Органы ротовой полости. Ротовая полость относится к переднему отделу пищеварительного канала. Ее эпителий имеет эктодермальное происхождение. Слизистая оболочка ее не имеет мышечной пластинки и местами отсутствует подслизистая основа. Мышечная оболочка представлена мышцами. Строение губы. Губы являются началом ротовой полости. Слизистая губы переходит с кожи, поэтому здесь погранично располагаются в толще губы: 1 - кожная часть или промежуточная, 2 - переходная часть (промежуточная), 3 - собственно слизистая оболочка (внутренняя часть). Кожная часть имеет строение кожи. В коже имеются волосы. Железы встречаются и сальные, и потовые. Переходная часть делится на: - наружную (гладкая) зону, - внутреннюю (ворсинчатая), сосочковая. Наружная зона – то, что находится за линией смыкания губ. Эпидермис тонкий, особенно роговой слой, поэтому капилляры просвечиваются легко, красный цвет. Соединительная ткань лежит под эпидермисом и не образует глубоких сосочков, гладко соприкасается с эпителием. Под эпидермисом–сальные железы, а потовые постепенно редуцируются. В области смыкания губ выделяют ворсинчатую часть. Эпителий здесь толстый. В него вдаются глубокие сосочки. В соединительной ткани расположены крупные кровеносные сосуды. Они обеспечивают окраску и необходимы для согревания или охлаждения пищи и др. Слизистая (внутренняя) часть губы представлена слизистой оболочкой (многослойный плоский неороговевающий эпителий). Собственная пластинка слизистой с кровеносными сосудами образуют переход в подслизисую основу, где располагаются секреторные отделы трубчатоальвеолярных слюнных желез, они довольно крупные. Глубже расположены мышцы губы – поперечно-полосатые мышцы, они еще недоразвиты. Пучки мышечных волокон: циркулярные и продольные. ЯЗЫК – производная стенки ротовой полости. Функции языка: 1. удержание пищи 2. механическая переработка пищи 3. функция речи 4. вкусовая рецепция. В языке различают 2 поверхности: 1- передняя (спинка), 2- задняя. Слизистая спинки языка имеет ряд особенностей. Выделяют 4 типа сосочков. По всей спинке лежат 1) нитевидные (конусовидные) сосочки диаметром 0,3 мм. Они покрыты многослойным плоским неорговевающим эпителием. 2) грибовидные сосочки (до 1,5 мм в высоту) у них более широкая шляпка и узкое основание. В их основании – соединительнотканный сосочек, от которого внутрь отходят 15-20 вторичных сосочков. 3) листовидные, от 4 до 8 пар по краю языка, встречаются только в детском возрасте (7-8 лет) и по боковой поверхности языка в виде двух групп по 6-7 шт. Листовидные сосочки имеют первичные соед.-тканные сосочки, состоящие из 2-3 сосочков. В эпителии расположены вкусовые луковицы в большом количестве, особенно на боковой поверхности. С возрастом они редуцируются и на этом месте формируется жировая ткань. 4) У основания языка на границе его корня – крупные желобоватые сосочки (6-12 шт), окруженные валом. Они не выдаются над поверхностью, а углублены, их толщина 3-4 мм. Эпителий содержит вкусовые луковицы на боковой части. Желоб окружает сосочек. В желоб при разжевывании пищи часть попадает для дегустации. В него под большим давлением выбрасывается слюна и вымывает его содержимое. Основу языка составляет поперечно-полосатая мышечная ткань, которая образует мышцы языка. Мышцы языка идут в 3-х направлениях: продольном, вертикальном и поперечном. 3. Большие слюнные железы. Классификация, источники развития, строение и функции. Эндокринная функция. Возрастные изменения. Все слюнные железы образуются в эмбриогенезе на 6-8 нед (к 2-м мес). Эти железы сложные альвеолярные или альв.трубчатые разветвленные, имеют дольчатость. Они имеют: 1-капсула; 2-междольковая соед.ткань, в которой лежат междольковые выводные протоки и кровеносные сосуды. Дольки представлены секреторными отделами и внутридольковыми выводными протоками. Внутридольковый выводной проток складывается из вставочных отделов, которые начинаются от секреторных отделов и из исчерченных протоков. Секреторные отделы в околоушной железе чисто белковые, т.е. серозные. Секрет жидкий (в основном–Н2О) и содержит большое кол-во ферментов (мальтаза, амилаза). В них выделяется инсулиноподобный фактор, фактор роста эпителия, фактор роста нервов. Секреторные отделы располагаются очень плотно по отношению друг к другу. Имеются также миоэпителиальные клетки, создающие многослойность и подчеркивающие эктодермальность происхождения железы. Вставочные протоки ветвящиеся, выстланы кубическим эпителием и миоэпителиальными клетками. Исчерченность выражена за счет складок цитолеммы и расположенных в складках митохондрий. Междольковые протоки выстланы многослойным эпителием, в месте выхода многослойным кубическим. Выводной проток открывается в ротовую полость на уровне 2-го верхнего большого коренного зуба. Поднижнечелюстная – сложная разветвленная альвеолярно-трубчатая, смешанная по характеру секрета. Содержит секреторные отделы белкового и белково-слизистого типа. Центр часть занимают светлые слизистые клетки, а по периферии – резко базофильные белковые клетки (полулунья Джиануцци). Снаружи их расположены миоэпителиальные клетки. Эти клетки в первый период вырабатывают слизистый секрет. Вставочные и исчерченные протоки более ветвящиеся. Выводные протоки могут располагаться группой (3-4). Железа, учитывая различный клеточный состав, окрашивается пестро, т.к. слизистые клетки более светлые, белковые – базофильные. Выводные протоки открываются в области уздечки языка (подле выводным протоком подъязычной железы). Подъязычная железа – смешанного характера (слизисто-белкового). Белковых секреторных отделов очень мало, и они быстро ослизняются. Смешанные секреторные отделы, где белковые клетки с возрастом подвержены ослизнению; в зрелом возрасте имеются только слизистые секреторные отделы. Считается, что подъязычная железа является чисто слизистой. Слюнные железы – околоушные, подчелюстные, подъязычные. Представляют собой сложные альвеолярные или альвеолярно – трубчатые железы. Экзокринная функция заключается в регулярном отделении в ротовую полость слюны. Экскреторная функция - со слюной во внешнюю среду выделяются различные органические и неорганические вещества: мочевая кислота, креатин, железо, йод и др. Защитная функция слюнных желез состоит в выделении бактерицидного вещества — лизоцима. Эндокринная функция слюнных желез обеспечивается наличием в слюне биологически активных веществ типа гормонов — инсулина, паротина, фактора роста эпителия (ФРЭ), фактора летальности и др. Слюнные железы активно участвуют в регуляции водно-солевого гомеостаза. 4. Зубы: строение и источники развития. Смена зубов. Возрастные изменения. Являются важной частью пищеварительного аппарата. Они участвуют в размельчении пищи. Большую роль играют в произношении звуков. Зубы образуются 2-мя генерациями: молочные и постоянные. Эмбриональные зачатки зуба возникают в конце 2- го месяца внутриутробного развития. В первую очередь во время разграничения вестибулярной части в ротовой полости образуется эпителиальный валик. От него вглубь тканей формируются эпителиальные выпячивания – зубная пластинка. На ней по количеству зубов образуются эпителиальные выросты, из которых образуются эмалевые органы. Таким образом, эмалевые органы имеют эпителиальное эктодермальное происхождение. Зубной сосочек подвергается своей дифференцировке под действием энамелобластов, их базальных мембран, при этом дифференцировка ускоряется. В наружном слое зубного сосочка дифференцируются клетки одонтобласты. Эти клетки имеют тело с органеллами, прилежащее к мезенхиме, а от тела в сторону эмали формируется отросток, который может иметь небольшие отверстия. Эти отростки идут строго радиально. Первые элементы солей (фосфор, кальций, фториды) начинают откладываться на границе с эмалью в наружной части дентина. Внутри зубного сосочка оставшиеся ткани формируют мякоть зуба (пульпу). В ней образуются кровеносные капилляры, нервные окончания и дифференцируются клетки, среди которых выделяется соединительная ткань пульпы и малодифференцированные клетки, из которых постепенно могут образовываться новые одонтобласты. Позднее, в области корня зуба и шейки, снаружи от дентина, начинает образовываться цемент, который по строению близок к костной ткани; он укрепляется в лунке челюсти при помощи периодонта. В конце 4 месяца эмбриогенеза на зубной пластинке начинают формироваться зачатки постоянных зубов и в течение долгого периода времени зачатки молочных зубов и зачатки постоянных зубов находятся в одной полости. Эмаль – плотная ткань, на 96-97% представлена неорганическими соединениями (соли кальция, фосфаты, фториды и др.). Из органических – коллаген, гликозаминогликаны, щелочная фосфатаза, протеогликаны (3-4%). На шлифе зуба в эмали выделяется чередование тѐмных и светлых полос. Дентин – плотная твѐрдая ткань. 70%–неорганических соединений (соли кальция, магния, фосфора, меньше фторидов). В нѐм выделяют: 1) предентин – более внутренний слой; 2) дентин – более наружный слой. Пульпа. Имеет сложное строение. В ней выделяют: центральную, промежуточную и наружную части. В центральной части располагаются тонкостенные капилляры, скопления макрофагов, фибробластов. В промежуточной – малодифференцированные элементы, из которых могут образовываться одонтобласты. В наружном слое – тела одонтобластов, проходят нервные волокна по дентинным канальцам к чувствительным окончаниям и до эмали. 5. Строение и тканевой состав стенки глотки и пищевода в различных его отделах. Железы пищевода, их гистофизиология. Пищевод — орган слоистого типа. Стенка образована 4-мя оболочками: слизистой, подслизистой, мышечной и адвентициальной (серозной). Слизистая оболочка образует продольные складки и состоит из трех слоев: эпителиального, собственной пластинки и мышечной пластинки. Эпителиальный слой — многослойный плоский неороговевающий эпителий, образованный базальным, шиповатым и слоем плоских клеток. Собственная пластинка слизистой оболочки образована рыхлой волокнистой соединительной тканью. Ее основные структуры — кровеносные и лимфатические сосуды, нервные волокна, одиночные лимфоидные фолликулы, выводные протоки собственных желез пищевода и концевые отделы кардиальных желез пищевода, которые встречаются только в двух местах: на уровне перстневидного хряща гортани и пятого хряща трахеи или в нижней части пищевода близ его входа в желудок. Это простые разветвленные трубчатые железы, похожие на кардиальные железы желудка, откуда их название. Концевые отделы состоят из кубических или цилиндрических мукоцитов, вырабатывающих слизь. Мышечная пластинка слизистой оболочки образована продольными пучками гладкой мышечной ткани. Она участвует в формировании складок, облегчает прохождение грубых комков пищи. Подслизистая оболочка образована рыхлой волокнистой соединительной тканью и участвует в образовании складок слизистой оболочки, обеспечивает ее питание и подвижность. Мышечная оболочка образована внутренним циркулярным и наружным продольным слоями. В верхней трети — поперечно-полосатой, в средней третии поперечно-полосатой, и гладкой, в нижней трети — только гладкой мышечной тканью. Циркулярный слой мышечной оболочки образует верхний и нижний сфинктеры пищевода. Функция оболочки — продвижение пищи к желудку. Между слоями мышечной оболочки находится межмышечное нервное сплетение Ауэрбаха. Серозная оболочка входит в состав стенки пищевода только в его поддиафрагмальном отделе. Образована двумя слоями: внутренний — рыхлая волокнистая соединительная ткань, наружный — мезотелий. На остальной части наружная оболочка представлена адвентицией, содержащей множество сосудов и нервное сплетение. Глотка - конусообразный канал длиной 12-14 см, соединяющий ротовую полость с пищеводом. Стенка глотки построена из трех оболочек - слизистой с подслизистой основой, мышечной и адвентициальным. Различают три части глотки - носовую, ротовую и гортанную. Слизистая оболочка носовой части глотки покрыта однослойным многорядным реснитчатым эпителием (респираторного типа). Слизистая оболочка ротовой и гортанной части глотки, подобно ротовой полости, выстлана многослойным плоским неороговевающим эпителием. Рыхлая соединительная ткань собственной пластинки слизистой оболочки сливается с подслизистой основой, где расположены концевые секреторные отделы слизистых желез глотки. Мышечная оболочка образована исполосовано мышечной тканью и формирует два слоя - наружный циркулярный и внутренний продольный. Адвентициальным оболочка образована рыхлой волокнистой соединительной тканью. 6. Строение стенки желудка. Строение слизистой оболочки в различных отделах органа. Макроскопически желудок состоит из 4-х отделов: кардиального, фундального, тела и пилорического. Гистологически же выделяют только три отдела, т. к. дно и тело желудка сходны по строению и расцениваются как один отдел. Все отделы имеют некоторые особенности гистологического строения слизистой оболочки, в частности, желудочных желез. Желудок — орган слоистого типа. Состоит из четырех оболочек: слизистой, подслизистой, мышечной и серозной. Слизистая оболочка имеет сложный рельеф, представленный желудочными ямками, складками и полями. Ямки — это углубления эпителия в собственную пластинку слизистой оболочки. Складки представляют собой выпячивания в просвет желудка слизистой и подслизистой оболочек. Поля — это участки слизистой оболочки, включающие группу желез, отграниченную от других таких же групп выраженной прослойкой рыхлой волокнистой соединительной ткани с просвечивающими кровеносными сосудами. Ямки и складки существенно увеличивают рабочую поверхность слизистой оболочки. Слизистая оболочка состоит из трех слоев: эпителиального, собственной и мышечной пластинок. Эпителиальный слой представлен однослойным цилиндрическим железистым эпителием. Он образован железистыми эпителиоцитами — мукоцитами, секретирующими слизь. Слизь формирует непрерывный слой толщиной до 0,5 мкм, являясь важным фактором защиты слизистой желудка. Собственная пластинка слизистой оболочки образована рыхлой волокнистой соединительной тканью. В ней находятся мелкие кровеносные и лимфатические сосуды, нервные стволики, лимфоидные узелки. Основными структурами собственной пластинки являются железы. Все железы желудка простые трубчатые разветвленные. Они открываются в желудочные ямки и состоят из трех частей: дна, тела и шейки. В зависимости от локализации железы делятся на кардиальные, главные или фундальные и пилоричекие. Мышечная пластинка слизистой оболочки состоит из трех слоев гладкой мышечной ткани: внутреннего и наружного циркулярных и среднего продольного. Функция обеспечение подвижности слизистой, участие в формировании ее рельефа. Подслизистая оболочка образована рыхлой волокнистой неоформленной соединительной тканью, содержит артериальное и венозное сплетения, ганглии подслизистого нервного сплетения Мейснера. В некоторых случаях здесь могут располагаться крупные лмфоидные фолликулы. Мышечная оболочка образована тремя слоями гладкой мышечной ткани: внутренний косой, средний циркулярный, наружный продольный. В пилорическом отделе желудка циркулярный слой достигает максимального развития, формируя пилорический сфинктер. Серозная оболочка образована двумя слоями: слоем рыхлой волокнистой неоформленной соединительной ткани и лежащим на нем мезотелием. 7. Тонкая кишка. Строение стенки, её тканевой состав. Гистофизиология системы «крипта-ворсинка». Тонкая кишка состоит из 3 частей: 1) двенадцатиперстной, 2) тощей; 3) подвздошной. Стенка тонкой кишки состоит из 4 оболочек: 1) слизистой, включающей слой эпителия, собственную пластинку и мышечную пластинку; 2) подслизистой основы: 3) мышечной оболочки, состоящей из внутреннего циркулярного и наружного продольного слоев гладких миоцитов; 4) серозной. Источники развития эпителия — кишечная энтодерма: рыхлой соединительной и гладкой мышечной тканей — мезенхима; мезотелия серозной оболочки — висцеральный листок спланхнотома. Строение. Рельеф (поверхность) слизистой оболочки представлен складками, ворсинками и криптами (простыми трубчатыми железами). Складки слизистой оболочки образованы слизистой оболочкой и подслизистой основой, имеют циркулярное направление; называются полулунными, или циркулярными. Ворсинки — это выпячивания слизистой оболочки, в состав которых входят рыхлая соединительная ткань собственной пластинки, гладкие миоциты мышечной пластинки и однослойный призматический (кишечный) эпителий, покрывающий ворсинки. Эпителий, покрывающий ворсинки, называется столбчатым. Собственная пластинка слизистой оболочки представлена рыхлой соединительной тканью, в которой содержится много ретикулярных волокон и ретикулоподобных клеток. Кроме того, в собственной пластинке находятся одиночные лимфатические узелки. Мышечная пластинка слизистой оболочки тонкой кишки состоит из 2 слоев гладких миоцитов: 1) внутреннего циркулярного и 2) наружного продольного. Между этими слоями имеется прослойка рыхлой соединительной ткани. Подслизистая основа состоит из рыхлой соединительной ткани, в которой имеются все сплетения: нервное, артериальное, венозное и лимфатическое. В подслизистой оболочке двенадцатиперстной кишки находятся сложные разветвленные трубчатые железы. Мышечная оболочка стенки тонкой кишки состоит из 2 слоев гладких миоцитов: 1) внутреннего циркулярного и 2) наружного продольного. Между этими слоями находится прослойка рыхлой соединительной ткани, в которой расположены 2 нервных сплетения: 1) мышечно-кишечное и 2) чувствительное мышечно-кишечное. Серозная оболочка тонкой кишки состоит из соединительнотканной основы, покрытой мезотелием. Дупликатура серозной оболочки образует брыжейку кишечника, которая прикрепляется к дорсальной стенке брюшной полости. 8. Толстая кишка. Строение стенки, её тканевой состав. Особенности строения слизистой оболочки. Толстый кишечник состоит из 3 отделов: -Слепая кишка с червеобразным отростком -Ободочная кишка (восходящая, поперечная, нисходящая, сигмовидная) -Прямая кишка Стенка толстого кишечника образована 4 оболочками: 1.Слизистая оболочка – поверхность слизистой оболочки увеличена благодаря постоянным полулунным складкам. Ворсинки отсутствуют, кишечные крипты глубже, чем в тонком кишечнике. Состоит из 3 слоёв: Эпителий – однослойный призматический, содержит клетки 4 типов: Призматически клетки – располагаются на поверхности слизистой оболочки и в криптах. Бокаловидные клетки – находятся в криптах и на поверхности слизистой оболочки. Недифференцированные клетки – лежат в глубине крипт, являются камбиальными элементами эпителия кишки. Эндокринные клетки – располагаются на дне крипт. 2.Собственная пластинка – состоит из рыхлой волокнистой соединительной ткани. Содержит капилляры и нервные волокна. Ретикулярные волокна имеют вид густой сети. В ней располагаются также одиночные лимфатические пузырьки. 3.Мышечная пластинка – состоит из 2 слоёв гладкомышечных клеток: -Наружный – продольный Внутренний – циркулярный 2.Подслизистая основа – образована рыхлой волокнистой соединительной тканью с большим количеством эластических волокон, часто содержит жировую ткань. В ней располагаются лимфатические узелки, элементы подслизистых нервного (Мейснера), венозного и лимфатического сплетений. 3.Мышечная оболочка – образована 2 слоями гладкой мышечной ткани: Наружный – продольный (имеет вид 3 лент, между которыми мышечная ткань развита слабо) Внутренний – циркулярный. Между слоями мышечной оболочки располагаются прослойки соединительной ткани и элементы межмышечного нервного сплетения (Ауэрбаха). 4.Серозная оболочка – покрывает одни отделы толстой кишки полностью, другие – частично, где замещается адвентицией. 9. Червеобразный отросток. Особенности строения и функции. Прямая кишка. Строение стенки. Червеобразный отросток (аппендикс) – пальцевидный вырост слепой кишки с узким звёздчатым или неправильной формы просветом, который содержит клеточный детрит и может облитерировться. Стенка его относительно толстая вследствие высокого содержания в ней лимфоидной ткани. Особенности строения: -Собственная пластинка содержит короткие крипты, а также многочисленные лимфатические узелки и межфолликулярные скопления лимфоидной ткани. -Мышечная пластинка развита слабо -Наружный (продольный) слой мышечной оболочки – сплошной -Серозная оболочка полностью покрывает червеобразный отросток Функции аппендикса: Червеобразный отросток выполняет защитную функцию и является периферическим органом иммунной системы. Он обеспечивает поглощение антигенного материала, его представление иммунокомпетентным клеткам с развитием иммунных реакций. Прямая кишка. Стенка прямой кишки (rectum) состоит из тех же оболочек, что и стенка ободочной кишки. В тазовой части прямой кишки ее слизистая оболочка имеет три поперечные складки. В образовании этих складок участвуют подслизистая основа и кольцевой слой мышечной оболочки. Ниже этих складок располагается 8—10 продольных складок, между которыми видны углубления. В анальной части кишки различают три зоны: столбчатую, промежуточную и кожную. Слизистая оболочка прямой кишки состоит из эпителия, собственной и мышечной пластинок. Эпителий в верхнем отделе прямой кишки однослойный призматический, в столбчатой зоне нижнего отдела — многослойный, кубический, в промежуточной — многослойный плоский неороговевающий, в кожной — многослойный плоский ороговевающий. Переход от многослойного кубического эпителия к многослойному плоскому совершается резко в виде зигзагообразной — аноректальной линии (linea anorectalis). Собственная пластинка слизистой принимает участие в формировании складок прямой кишки. Здесь располагаются одиночные лимфоидные узелки и сосуды. В кожной зоне, окружающей анальное отверстие, к сальным железам присоединяются волосы. Потовые железы в собственной пластинке слизистой оболочки появляются на расстоянии 1 – 1,5 см от ануса, представляют собой трубчатые железы, концевые отделы которых свернуты кольцом (gll. circumanales). Это железы апокринового типа, в секрете которых обнаруживаются феромоны. Мышечная пластинка слизистой оболочки, как и в других отделах толстой кишки, состоит из двух слоев. Пучки гладких миоцитов ее постепенно переходят в узкие продольные пучки, тянущиеся до столбчатой зоны. В подслизистой основе прямой кишки располагаются сосудистые и нервные сплетения. Здесь же встречаются чувствительные пластинчатые нервные тельца. Мышечная оболочка прямой кишки состоит из двух слоев: внутреннего — циркулярного и наружного – продольного. Циркулярный слой на разных уровнях прямой кишки образует два утолщения, которые выделяются как отдельные анатомические образования — внутренний и наружный сфинктеры (m. sphincter ani internus et externus). Последняя мышца в отличие от всей мышечной оболочки образована поперечнополосатой мышечной тканью. Наружный продольный слой мышечной оболочки прямой кишки в противоположность другим отделам толстой кишки сплошной. Между обоими мышечными слоями имеется прослойка рыхлой волокнистой неоформленной соединительной ткани, в которой лежат мышечно-кишечные сплетения и сосуды. Серозная оболочка покрывает прямую кишку в верхней ее части; в нижних отделах прямая кишка имеет соединительнотканную оболочку. 10. Поджелудочная железа. Строение экзо- и эндокринного отделов. Цитофизиологическая характеристика ацинарных клеток. Типы эндокриноцитов, их морфофункциональная характеристика. Поджелудочная железа является смешанной железой, включающей экзокринную и эндокринную части. В экзокринной части вырабатывается панкреатический сок, богатый пищеварительными ферментами — трипсином, липазой, амилазой, поступающий по выводному протоку в двенадцатиперстную кишку, где его ферменты участвуют в расщеплении белков, жиров и углеводов до конечных продуктов. В эндокринной части синтезируется ряд гормонов — инсулин, глюкагон, соматостатин, принимающие участие в регуляции углеводного, белкового и жирового обмена в тканях. Развитие. Поджелудочная железа развивается из энтодермы и мезенхимы. Ее зачаток появляется в конце 3-й недели эмбриогенеза. Начинается дифференцировка на экзокринные и эндокринные отделы железы. В экзокринных отделах образуются ацинусы и выводные протоки, а эндокринные отделы превращаются в островки. Из мезенхимы развиваются соединительнотканные элементы стромы, а также сосуды. Строение. Поджелудочная железа с поверхности покрыта тонкой соединительнотканной капсулой, срастающейся с висцеральным листком брюшины. Ее паренхима разделена на дольки, между которыми проходят соединительнотканные тяжи. В них расположены кровеносные сосуды, нервы, интрамураль-ные нервные ганглии, пластинчатые тельца и выводные протоки. Экзокринная часть: представлена панкреатическими ацинусами, вставочными и внутридольковыми протоками, а также междольковыми протоками и общим панкреатическим протоком, открывающимся в двенадцатиперстную кишку. Эндокринная часть представлена панкреатическими островками, островками Лангерганса, лежащими между панкреатическими ацинусами. Островки состоят из инсулоцитов. Возрастные изменения. Изменяются соотношения между ее экзокринной и эндокринной частями. Островки наиболее сильно развиты в железе в первые годы жизни. С возрастом их число постепенно уменьшается. Регенерация. Пролиферативная активность клеток поджелудочной железы крайне низкая, поэтому в физиологических условиях в ней происходит обновление клеток путем внутриклеточной регенерации. 11. Печень. Особенности строения, кровоснабжения, функции. Печень состоит из правой и левой долей, покрыта соединительнотканной капсулой, от которой отходят прослойки соединительной ткани. Развитие. Начинается на 5-й неделе эмбриогенеза в виде выпячивания стенки энтодермальной кишки. Это выпячивание называется печеночной бухтой. Из краниальной части печеночной бухты развиваются правая и левая доли печени и ее протоки, из каудальной — желчный пузырь и его проток. Из устья бухты формируется общий желчный проток, который открывается в двенадцатиперстную кишку. Строма печени развивается из мезенхимы. Общий план строения. Печень снаружи покрыта соединительнотканной капсулой (Глиссоновой капсулой). Капсула покрыта брюшиной, прочно срастающейся с капсулой. Паренхима печени состоит из 500 ООО печеночных долек. Печеночная долька — структурно-функциональная единица печени, имеет форму призмы, расширенной у основания и суженной у вершины. Простые дольки могут сливаться, образуя сложные дольки. У человека дольки печени отделены друг от друга тонкими прослойками соединительной ткани. В междольковых прослойках соединительной ткани (строме печени) проходят междольковые и вокругдольковые кровеносные сосуды и желчные протоки, и нервы. Кровеносная система печени. Кровеносная система печени отличается тем, что в ворота печени входят печеночная артерия, приносящая артериальную кровь, и воротная вена, несущая венозную кровь от непарных органов брюшной полости. Каждый из этих сосудов разветвляется на долевые - сегментарные - междольковые - вокругдольковые капилляры. Капилляр, отходящий от вокругдольковой вены, сливается с капилляром, отходящим от вокругдольковой артерии. В результате этого слияния образуется внутридольковый печеночный капилляр, по которому течет смешанная (артериальная и венозная) кровь. Внутридольковые капилляры впадают в центральную вену дольки, по которой кровь поступает в поддольковую вену. Поддольковые вены впадают в ветви печеночных вен. Печеночные вены в количестве 3-4 впадают в нижнюю полую вену. Классические печеночные дольки состоят из печеночных балок. Балки состоят из печеночных клеток. или гепатоцитов. Между печеночными балками проходят внутридольковые синусоидные капилляры. Балки и капилляры расположены радиально. 12. Желчный пузырь и желчевыводящие пути. Строение и функции. Желчный пузырь (vesica biliaris seu fellae) представляет собой тонкостенный орган (толщина стенки 1,5-2 мм), вмещающий 40-70 мл желчи. Стенка желчного пузыря состоит из трех оболочек: слизистой, мышечной и адвентициальной (рис. 16.42). Желчный пузырь со стороны брюшной полости покрыт серозной оболочкой. Слизистая оболочка образует многочисленные складки. Она выстлана однослойным столбчатым эпителием. В составе эпителия различают поверхностные эпителиоциты, бокаловидные клетки и базальные (камбиальные) клетки. Поверхностные столбчатые эпителиоциты (холецистоциты) имеют микроворсинчатую каемку. Под эпителием располагается собственная пластинка слизистой оболочки, содержащая большое количество эластических волокон. Там же встречаются жировые, тучные и плазматические клетки. В области шейки пузыря в ней находятся альвеолярно-трубчатые железы, выделяющие слизь. Эпителий слизистой оболочки обладает способностью всасывать воду и некоторые другие вещества из желчи, заполняющей полость пузыря. Поэтому пузырная желчь всегда более густой консистенции и более темного цвета, чем желчь, изливающаяся непосредственно из печени. Мышечная оболочка желчного пузыря состоит из пучков гладких миоцитов, расположенных в виде сети, в которой миоциты располагаются преимущественно циркулярно. В области шейки пузыря циркулярные пучки мышечных клеток особенно сильно развиты. Вместе с мышечным слоем пузырного протока они образуют сфинктер. Между пучками мышечных клеток всегда имеются хорошо выраженные прослойки рыхлой соединительной ткани. Адвентициальная оболочка желчного пузыря состоит из плотной соединительной ткани, в которой содержится много толстых эластических волокон, образующих сети. Со стороны брюшной полости стенку желчного пузыря покрывает серозная оболочка. XV. ДЫХАТЕЛЬНАЯ СИСТЕМА 1. Внелегочные воздухоносные пути (носовая полость, гортань, трахея). Особенности строения стенки воздухоносных путей. Тканевой состав и морфофункциональная характеристика их оболочек. Клеточный состав эпителия слизистой оболочки. Дыхательная система — это совокупность органов, обеспечивающих в организме внешнее дыхание, а также ряд важных недыхательных функций. В состав дыхательной системы входят различные органы, выполняющие воздухопроводящую и дыхательную (газообменную) функции: полость носа, носоглотка, гортань, трахея, внелегочные бронхи и легкие. Развитие. Гортань, трахея и легкие развиваются из одного общего зачатка, который появляется на 3—4-й неделе путем выпячивания вентральной стенки передней кишки. Из мезенхимы, окружающей растущее бронхиальное дерево, дифференцируются гладкая мышечная ткань, хрящевая ткань, волокнистая соединительная ткань бронхов, эластические, коллагеновые элементы альвеол, а также прослойки соединительной ткани, прорастающие между дольками легкого. Носовая полость. В носовой полости различают преддверие и собственно носовую полость, включающую дыхательную и обонятельную области. Строение. Преддверие образовано полостью, расположенной под хрящевой частью носа. Оно выстлано многослойным плоским ороговевающим эпителием. Под эпителием в соединительнотканном слое заложены сальные железы и корни щетинковых волос. В более глубоких частях преддверия эпителий становится неороговевающим, переходящим в многорядный, реснитчатый. Внутренняя поверхность собственно носовой полости вдыхательной части покрыта слизистой оболочкой, состоящей из многорядного призматического реснитчатого эпителия и соединительнотканной собственной пластинки, соединенной с надхрящницей или надкостницей. В эпителии, расположенном на базальной мембране, различают 4 вида клеток: реснитчатые, микроворсинчатые, базальные и бокаловидные. Гортань — орган воздухоносного отдела дыхательной системы, принимающий участие не только в проведении воздуха, но и в звукообразовании. Гортань имеет три оболочки: слизистую, фиброзно-хрящевую и адвентициальную. Слизистая оболочка выстлана многорядным реснитчатым эпителием. Собственная пластинка слизистой оболочки, представленная рыхлой волокнистой соединительной тканью, содержит многочисленные эластические волокна. На передней поверхности в собственной пластинке слизистой оболочки гортани содержатся смешанные белковослизистые железы. Здесь же имеются значительные скопления лимфатических узелков, носящие название гортанных миндалин. В средней части гортани имеются складки слизистой оболочки, образующие так называемые истинные и ложные голосовые связки. В слизистой оболочке выше и ниже истинных голосовых связок располагаются смешанные белковослизистые железы. Фиброзно-хрящевая оболочка состоит из гиалиновых и эластических хрящей, окруженных плотной волокнистой соединительной тканью. Она выполняет роль защитно-опорного каркаса гортани. Адвентициальная оболочка состоит из коллагеновой соединительной ткани. Трахея — полый трубчатый орган, состоящий из слизистой оболочки, подслизистой основы, волокнисто-хрящевой и адвентициальной оболочек. Слизистая оболочка при помощи тонкой подслизистой основы связана с фибрознохрящевой оболочкой трахеи и благодаря этому не образует складок. Она выстлана многорядным призматическим реснитчатым эпителием, в котором различают реснитчатые, бокаловидные, эндокринные и базальные клетки. Подслизистая основа трахеи состоит из рыхлой волокнистой соединительной ткани, переходящей в плотную волокнистую соединительную ткань надхрящницы незамкнутых хрящевых колец. В подслизистой основе располагаются смешанные белково-слизистые железы. Волокнисто-хрящевая оболочка трахеи состоит из гиалиновых хрящевых колец, не замкнутых на задней стенке трахеи. Адвентициальная оболочка трахеи состоит из рыхлой волокнистой неоформленной соединительной ткани, которая соединяет этот орган с прилежащими частями средостения. Бронхи крупного калибра характеризуются складчатой слизистой оболочкой, благодаря сокращению гладкой мышечной ткани, многорядным реснитчатым эпителием, наличием желез, крупных хрящевых пластин в фибрознохрящевой оболочке. Бронхи среднего калибра отличаются меньшей высотой клеток эпителиального пласта и снижением толщины слизистой оболочки, наличием желез, уменьшением размеров хрящевых островков. В бронхах малого калибра эпителий реснитчатый двухрядный, а затем однорядный, хряща и желез нет, мышечная пластинка слизистой оболочки становится более мощной по отношению к толщине всей стенки. 2. Легкие. Внутрилегочные воздухоносные пути: бронхи и бронхиолы, строение их стенок в зависимости от их калибра. Плевра. Морфофункциональная характеристика. Легкое - парный комплексный орган, выполняющий целый ряд дыхательных и недыхательных функций Снаружи легкое покрыто серозной оболочкой - висцеральной плеврой. Она состоит из соединительнотканной пластинки и мезотелия(однослойный плоский эпителий). Легкое. Это бронхиальное дерево и респираторный отдел. Бронхиальное дерево относится к воздухоносным путям легких. Оно начинается главными бронхами крупного калибра (диаметр — около 15 мм), отходящими от трахеи (бифуркация трахеи). От главных бронхов отходят по 2 внелегочных долевых бронха 1-го порядка крупного калибра (диаметр — около 12 мм). От этих бронхов отходят 4 внелегочных зональных бронха 2-го порядка крупного калибра (диаметр 10-6 мм). От бронхов 2-го порядка отходят 10 внутрилегочных сегментарных бронха 3-го порядка среднего калибра (диаметр — около 5 мм). От них отходят субсегментарные бронхи 4-го порядка среднего калибра (диаметр 4-3 мм), которые переходят в субсегментарные бронхи 5-го порядка среднего калибра (диаметр 3 мм). От бронхов 5-го порядка отходят бронхи малого калибра, или малые бронхи (диаметр 2-1 мм). Малые бронхи разветвляются на терминальные (конечные) бронхиолы, диаметр которых составляет 1-0,5 мм. Этими бронхиолами заканчивается бронхиальное дерево. Строение стенки бронхов крупного и среднего калибров. Стенка бронхов этих калибров включает 4 оболочки: 1) слизистую; 2) подслизистую основу: 3) фиброзно-хрящевую: 4) адвентициальную. Строение стенки бронхов малого калибра. Стенка бронхов этого калибра вклучает 2 оболочки: 1( слизистую и 2) адвентициальную. Терминальные бронхиолы. Стенка терминальных бронхиол состоит из 2 истонченных оболочек: 1) слизистой и 2) адвентициальной. Слизистая оболочка состоит из 3 слоев: 1) эпителиальной пластинки. 2) собственной пластинки и 3) мышечной пластинки. Эпителиальная пластинка представлена кубическим реснитчатым эпителием, среди клеток которого имеются секреторные клетки Клара, каемчатые и безресиитчатые клетки. Соединительнотканная пластинка (РВСТ + сосуды + нервный апп.) плотно прирастает к легкому. Мезотелий покрывает ее снаружи и выстилает плевральную полость. Эта полость находится между висцеральной и париетальной плеврой. Париетальная плевра имеет аналогичное с висцеральной строение. Ее соединительнотканная пластинка прирастает к тканям внутренней поверхности грудной клетки, а мезотелий выстилает плевральную полость. Плевральная полость – щелевидное пространство между висцеральной и париетальной плеврами. Оно заполнено небольшим количеством плевральной жидкости, которая является продуктом секреции мезотелия и фильтрации плазмы крови. 3. Респираторные отделы легких. Аэрогематический барьер. Особенности кровоснабжения легкого. Респираторный отдел легких представлен совокупностью ацинусов. Ацинус – это структурно-функциональная единица респираторного отдела легких. В обоих легких начитывается до 150 тысяч ацинусов. 12 – 18 ацинусов составляют легочную дольку. Ацинусы выполняют функцию газообмена между вдыхаемым атмосферным воздухом и кровью в капиллярах малого круга кровообращения. Каждый из ацинусов представлен респираторными бронхиолами, альвеолярными ходами, альвеолярными мешочками и альвеолам. Внутриорганное кровоснабжение легких. В легких имеются разветвления кровеносных сосудов малого и большого круга кровообращение. ● Сосуды малого круга относятся к функциональной системе внутриорганного кровообращения. Ее гемокапилляры обеспечивают газообмен между кровью и атмосферным воздухом. Этот процесс называется «внешним дыханием». Он заключается в насыщении крови кислородом и выведении из нее углекислого газа. ●Сосуды большого круга относятся к трофической системе внутриорганного кровообращения. Ее гемокапилляры обеспечивают трофику тканей легкого. Аэрогематический барьер. Между атмосферным вдыхаемым воздухом в альвеолах и кровью в гемокапиллярах малого круга кровообращения имеется аэрогематический барьер. Он определяет режим обмена кислорода и углекислого газа по векторам их парциальных давлений между кровью и атмосферным воздухом. Этот барьер также препятствует проникновению в кровь из вдыхаемого воздуха большинства бактерий и микропылевых частиц. Этот барьер проницаем для вирусов, угарного газа, боевых отравляющих газов, продуктов возгонки никотина, наркотических газообразных веществ и паров. XVI. К О Ж А И ЕЁ П Р О И З В О Д Н Ы Е. 1. Кожа. Эпидермис. Слои эпидермиса. Основные диффероны клеток в эпидермисе. Процесс кератинизации кожи, его значение. Общий план строения кожи. Кожа состоит из трех основных частей: эпидермиса, дермы и гиподермы (рис. 1), которые имеют различные источники эмбрионального происхождения. В составе кожи имеются также структуры производные эпидермиса:кожные железы (потовые, сальные, молочные), волосы и ногти. По особенностям строения выделяют два типа кожи: толстую (ладони и подошвы) и тонкую (остальные участки тела). Строение основных частей кожи. Эпидермис, дерма и гиподерма имеют различное тканевое представительство. Эпидермис – наружная часть кожи, непосредственно соприкасающаяся с внешней средой. Первый эпидермис (однослойный) развивается их кожной эктодермы в конце третьей недели эмбриональной жизни. Эктодермальный многослойный плоский ороговевающий эпителий, представленный кератиноцитами, является структурной основой эпидермиса. Эпителиоциты-кератиноциты расположены в эпидермисе пятью слоями. Выделяют базальный, шиповатый, зернистый, блестящий и роговой слои. После разрушения межклеточных контактов кератиноциты перемещаются вверх из слоя в слой. В эпидермисе толстой кожи развиты все пять слоев. В эпидермисе тонкой кожи слабо выражен зернистый слой, нет блестящего слоя, роговой слой тонкий. Кератиноциты базального слоя лежат на базальной мембране. Среди них расположены немногочисленный стволовые клетки. Дерма – соединительнотканная основа кожи. Толщина дермы толстой кожи достигает 3 – 5 мм, тонкой - 0,5 - 2мм (на спине до 7мм). У мужчин дерма толще, чем у женщин. В дерме находятся волосяные фолликулы и секреторные (концевые) отделы кожных желез (см.далее). Дерма состоит из двух слоёв: сосочкового(волокнистой неоформленной соединительной тканью) и сетчатого (плотной волокнистой неоформленной соединительной ткани). Гиподерма состоит из жировой ткани. Жировая ткань гиподермы тонкими прослойками соединительной ткани разделена на микродольки. С возрастом, а также при недостатке женских половых гормонов - эстрогенов прослойки соединительной ткани утолщаются и кожа может приобрести бугристый вид (целлюлит). 2. Производные кожи. Железы. Сальные и потовые железы (меро- и апокриновые), их развитие, строение, гистофизиология. САЛЬНЫЕ ЖЕЛЕЗЫ – экзокринные простые альвеолярные железы голокринового типа секреции. Их эмбриональное развитие тесно связано с формированием волоса, поэтому они присутствуют только на волосистой части кожи. На ладонях и подошвах их нет. - Секреторные (концевые) отделы сальных желез лежат на границе сосочкового и сетчатого слоев дермы. Они состоят из трех видов клеток: экзокриноцитов (секреторные клетки), миоэпителиоцитов (сократительные клетки, способствуют выдавливанию секрета) и камбиальных малодифференцированных клеток (обеспечивают регенерацию). Все они лежат на общей базальной мембране Выводные протоки желез открываются в волосяные воронки волосяных фолликулов Экзокриноциты желез синтезируют и накапливают в своей цитоплазме липиды. Вблизи выводного протока происходит разрушение секретирующих клеток. Их структурные фрагменты смешиваются с липидами и происходит окончательное формирование секрета железы – кожного сала. Функциональная активность сальных желез преимущественно регулируется половыми гормонами. ПОТОВЫЕ ЖЕЛЕЗЫ подразделяются на апокриновые (в подмышечных областях, в больших половых губах, вокруг ануса) и мерокриновые (в остальных областях тела). Это простые трубчатые железы. - Секреторные отделы находятся в сетчатом слое дермы. Они спирально закручены и имеют вид клубочков. В них различают: экзокриноциты (секреторные клетки), миоэпителиоциты (сократительные клетки) и камбиальные (малодифференцированные) клетки. Все клетки лежат на общей базальной мембране. -Выводные протоки открываются на поверхность эпидермиса. Железы имеются и в тонкой, и толстой коже. Мерокриновые железы иннервируются симпатической нервной системой. Функция апокриновых желез регулируется преимущественно половыми гормонами. За сутки взрослый человек выделяет 500-600 мл пота. Это количество может увеличиваться до 10 литров и более при тяжелой физической работе, банных процедурах, лихорадочных состояниях и пр. благодаря преимущественной активизации мерокриновых потовых желез. 3. Кожа. Дерма. Сосочковый и сетчатый слои, их тканевый состав. Гиподерма. Дерма – соединительнотканная основа кожи. Толщина дермы толстой кожи достигает 3 – 5 мм, тонкой - 0,5 - 2мм (на спине до 7мм). У мужчин дерма толще, чем у женщин. В дерме находятся волосяные фолликулы и секреторные (концевые) отделы кожных желез (см.далее). Дерма состоит из двух слоёв: сосочкового и сетчатого. Сосочковый слой находится непосредственно под эпидермисом. Он представлен рыхлой волокнистой неоформленной соединительной тканью. Сосочковый слой вдается сосочками в эпидермис прогибая базальную мембрану и прикрепляется к ней «якорными» коллагеновыми волокнами. В сосочковом слое расположены поверхностные кровеносные сосудистые сплетения, с большим количеством артериоло-венулярных анастомозов. Здесь же расположены поверхностные нервные сплетения кожи. Для сосочкового слоя характерно обилие рецепторных нервных окончаний: осязательные тельца Мейснера (осязание), колбы Краузе (воспринимают холодовые раздражения), тельца Руфини (воспринимают тепловые раздражения). Сетчатый слой расположен в глубине дермы и граничит с гиподермой. Он состоит из плотной волокнистой неоформленной соединительной ткани и создает механическую прочность кожи. В нем находятся глубокие сосудистые и нервные сплетения, которые анастомозируют с поверхностными. В сетчатом слое встречаются рецепторные нервные окончания – тельца Фатер-Пачини (воспринимают раздражения давлением и вибрацией). Гиподерма состоит из жировой ткани. Жировая ткань гиподермы тонкими прослойками соединительной ткани разделена на микродольки. С возрастом, а также при недостатке женских половых гормонов - эстрогенов прослойки соединительной ткани утолщаются и кожа может приобрести бугристый вид (целлюлит). Гиподерма прочно связана пучками коллагеновых волокон с сетчатым слоем кожи. Она способствует некоторой подвижности кожи по отношению к нижележащим тканям. Гиподерма является теплоизолятором, а также депо жира и связанной воды. У новорожденных детей в составе гиподермы имеется бурая жировая ткань. Её особенно много в межлопаточных областях. Она активно участвует в энергетическом и тепловом обмене. Её термогенез резко возрастает при охлаждениях организма. 4. Производные кожи. Волосы. Развитие, строение, рост и смена волос, иннервация. Ногти. Развитие, строение и рост ногтей. ВОЛОСЫ являются производными эпидермиса тонкой кожи. Различают длинные волосы (головы, бороды, усов), щетинистые (бровей, ресниц) и пушковые (на всех остальных частях тела). Волосы отсутствуют на подошвах и ладонях, т.е. в толстой коже. Волосы, образовавшиеся в период полового созревания, по характеру строения являются окончательными. В дальнейшем они подвергаются периодической смене. Жизнь волоса составляет в среднем от 2 до 5 лет. В волосе различают две основные части: волосяной фолликул и собственно волос. Собственно волос является продуктом ороговения кератиноцитов волосяного фолликула. Волосяной фолликул представляет собой пробиркообразную инвагинацию эпидермиса в дерму. Он состоит из соединительнотканной наружной волосяной сумки и двух (наружного и внутреннего) эпителиальных корневых влагалищ. Собственно волос представлен склеенными цементирующим аморфным матриксом роговыми чешуйками. Он состоит из стержня и корня. - Стержень свободно располагается над поверхностью кожи. - Корень находится в дерме волоса и заканчивается в ее сетчатом слое расширением. С этим расширением сливаются оба корневых эпителиальных влагалища. Таким образом, формируется луковица волоса. Волосяная соединительнотканная сумка плотно облегает луковицу и вдается в неё снизу в виде волосяного сосочка. В соединительной ткани сосочка много кровеносных капилляров, благодаря которым осуществляется трофика волоса. Волосяная луковица является матрицей нового волоса при замене выпавшего. НОГТИ – производные эпидермиса, характерные только для человека и высших приматов. Каждый ноготь имеет две основные части: ногтевую пластинку (собственно ноготь) и ногтевое ложе. Ногтевая пластинка является продуктом ороговения кератиноцитов ногтевого ложа. Ногтевое ложе состоит из следующих частей: - подногтевая пластинка (ростковый слой эпидермиса на базальной мембране), - задний и боковые ногтевые валики (кожные складки с ростковым и ороговевающим слоями эпидермиса), - ногтевые щели (щелевидные пространства между валиками и подногтевой пластинкой), - надногтевая пластинка (роговой слой заднего ногтевого валика, нарастающий на корень ногтевой пластинки, через него в области основания ногтя просвечивает светлый участок корня полулунной формы – луночка ногтя), - ногтевая матрица (участок эпителия подногтевой пластинки, на которой лежит корень, обеспечивает рост ногтя со скоростью около 1 мм в сутки). Ногтевая пластинка состоит из корня (лежит в задней ногтевой щели) и тела (состоит из спрессованных роговых чешуек с твердым кератином). 5. Возрастные особенности кожи детей. Кожа ребенка состоит из двух слоев: эпидермиса и дермы. Толщина эпидермиса у новорожденного и детей раннего возраста невелика и составляет приблизительно 0,15 — 0,25 мм, а у взрослого человека — от 0,25 до 0,35 мм. Три слоя эпидермиса у детей имеют ряд особенностей. Базальный слой у новорожденных выражен хорошо и представлен двумя видами клеток: базальными и меланоцитами. У новорожденных детей белой расы вследствие недоразвития меланоцитов кожа светлая, у новорожденных африканцев — более темная, красноватого оттенка. Чем меньше ребенок, тем зернистый слой кожи менее выражен, поэтому кожа у новорожденного прозрачная. Сквозь нее видны капилляры (через увеличительное стекло, микроскоп). Роговые придатки кожи (ногти, волосяной покров, сальные и потовые железы) продолжают развиваться после рождения. Например, волосы у новорожденных в основном пушковые, затем они выпадают, и начинается рост постоянных волос. На голове они растут медленно, с возрастом становятся толще, в период полового созревания появляются в подмышечных впадинах и на лобке. Ногти у доношенных новорожденных достигают концов фаланг пальцев, что является одним из признаков зрелости плода. Ногтевая пластинка очень чувствительна к различным неблагоприятным факторам. Кожа ребенка является также и сложным органом чувств. В ней заложено много рецепторов, воспринимающих раздражения внешней среды. У новорожденного и ребенка первых месяцев жизни кожа играет большую роль в приспособлении к факторам внешней среды. Кожа является также местом образования биологически активных веществ, необходимых для осуществления различных функций. XVII. МОЧЕВАЯ СИСТЕМА 1. Почки. Корковое и мозговое вещество почки. Нефрон – как морфофункциональная единица почки, его строение. Корковые и юкстамедуллярные нефроны. К мочевым органам относятся почки, мочеточники, мочевой пузырь и мочеиспускательный канал. Среди них почки являются мочеобразующими органами, а остальные составляют мочевыводящие пути. Развитие. В течение эмбрионального периода закладываются последовательно три парных выделительных органа: передняя почка, или предпочка, первичная почка и постоянная, или окончательная, почка. Предпочка образуется из передних 8—10 сегментных ножек мезодермы. Первичная почка формируется из большого числа сегментных ножек, расположенных в области туловища зародыша. Окончательная почка закладывается у зародыша на 2-м месяце, но развитие ее заканчивается лишь после рождения ребенка. Эта почка образуется из двух источников — мезонефрального протока и нефрогенной ткани. Почка — парный орган, в котором непрерывно образуется моча. Почки регулируют водно-солевой обмен между кровью и тканями, поддерживают кислотно-щелочное равновесие в организме, выполняют эндокринные функции. Строение. Почка покрыта соединительнотканной капсулой и, кроме того, спереди серозной оболочкой. Вещество почки подразделяется на корковое и мозговое. Корковое вещество располагается общим слоем под капсулой. Мозговое вещество разделено на 8—12 пирамид. Вершины пирамид, или сосочки, свободно выступают в почечные чашки. Мозговое вещество тонкими лучами врастает в корковое, образуя мозговые лучи. Строму почки составляет рыхлая волокнистая соединительная, интерстициальная ткань. Паренхима почки представлена эпителиальными почечными канальцами, которые при участии кровеносных капилляров образуют нефроны. Нефрон — структурная и функциональная единица почки. В состав нефрона входят капсула клубочка, проксимальный извитой каналец, проксимальный прямой каналец, тонкий каналец, в котором различают нисходящую часть и восходящую часть, дистальный прямой каналец и дистальный извитой каналец. Тонкий каналец и дистальный прямой каналец образуют петлю нефрона. Почечное тельце включает сосудистый клубочек и охватывающую его капсулу клубочка. Возрастные изменения. Особенно резким изменениям с возрастом подвергается длина нефронов. Их рост продолжается вплоть до половой зрелости. Поэтому с возрастом, по мере того как увеличивается масса канальцев, количество клубочков на единицу поверхности почки уменьшается. 2. Почки. Васкуляризация почки – кортикальная и юкстамедуллярная системы кровоснабжения. Морфофункциональные основы регуляции процесса мочеобразования. Васкуляризация. Кровь поступает к почкам по почечным артериям, которые, войдя в почки, распадаются на междолевые артерии. На границе между корковым и мозговым веществом они разветвляются на дуговые артерии. От них в корковое вещество отходят междольковые артерии. От междольковых артерии в стороны расходятся внутридольковые артерии, от которых начинаются приносящие артериолы. От верхних внутридольковых артерий приносящие артериолы направляются к коротким и промежуточным нефронам, от нижних — к юкстамедуллярным нефронам. В связи с этим в почках условно различают кортикальное кровообращение и юкстамедуллярное кровообращение. В кортикальной системе кровообращения приносящие артериолы распадаются на капилляры, образующие сосудистые клубочки почечных телец нефронов. Капилляры клубочков собираются в выносящие артериолы. Происходит первая фаза мочеобразования - процесс фильтрации жидкости и веществ из плазмы крови в нефрон. Выносящие артериолы вновь распадаются на капилляры, оплетающие канальцы нефрона и образующие перитубулярную капиллярную сеть. Происходит вторая фаза мочеобразования - процесс обратного всасывания части жидкости и веществ из нефрона в кровь. Из капилляров кровь перитубулярной сети собирается в верхних отделах коркового вещества сначала в звездчатые вены, а затем в междольковые. Последние впадают в дуговые вены, переходящие в междолевые, которые образуют почечные вены, выходящие из ворот почек. В юкстамедуллярной системе кровообращения выносящие клубочковые артериолы юкстамедуллярных нефронов идут в мозговое вещество, распадаясь на пучки тонкостенных сосудов, несколько более крупных, чем обычные капилляры, — прямые сосуды. В мозговом веществе как от выносящих артериол, так и от прямых сосудов отходят ветви для формирования мозговой перитубулярной капиллярной сети. Прямые сосуды образуют петли на различных уровнях мозгового вещества, поворачивая обратно. Нисходящие и восходящие части этих петель образуют противоточную систему сосудов, называемую сосудистым пучком. Капилляры мозгового вещества собираются в прямые вены, впадающие в дуговые вены. Возрастные изменения. Особенно резким изменениям с возрастом подвергается длина нефронов. Их рост продолжается вплоть до половой зрелости. Поэтому с возрастом, по мере того как увеличивается масса канальцев, количество клубочков на единицу поверхности почки уменьшается. 3. Мочевыводящие пути. Строение мочеточников, мочевого пузыря, мочеиспускательного и мочеполового каналов. К мочевым органам относятся почки, мочеточники, мочевой пузырь и мочеиспускательный канал. Среди них почки являются мочеобразующими органами, а остальные составляют мочевыводящие пути. Мочеточники: Слизистая оболочка - переходный эпителий. Включает 3 слоя клеток: базальный, промежуточный и поверхностный; Обладают выраженной способностью к растяжению – наличие глубоких продольных складок. Собственная пластинка слизистой оболочки –РВСт. Подслизистая основа – РВСт, в нижней половине мочеточников в подслизистой основе встречаются мелкие альвеолярно- трубчатые железы. Мышечная оболочка - Мышечная оболочка образована пучками гладких миоцитов (разделённых соединительнотканными прослойками) и содержит 2 или 3 слоя. В мочевых путях до середины мочеточников - 2 слоя: внутренний и наружный. С середины мочеточников и в пузыре - 3 слоя: внутренний, средний, наружный. Спиральная ориентация гладких миоцитов - порционный транспорт мочи. Мочеточник состоит из 3 цистоидов, между которыми сфинктеры. Наружная оболочка - является адвентициальной, т.е образована соединительной тканью. Мочевой пузырь: Слизистая оболочка - переходный эпителий. Включает 3 слоя клеток: базальный, промежуточный и поверхностный; Собственная пластинка слизистой оболочки –РВСт. Слизистая оболочка пустого пузыря образует много складок - кроме треугольной области у места впадения мочеточников. Подслизистая основа – РВСт, в области вышеуказанного треугольника в пузыре подслизистой основы нет (не образуются складки) Мышечная оболочка Мышечная оболочка образована пучками гладких миоцитов (разделённых соединительнотканными прослойками) и содержит 2 или 3 слоя. В мочевых путях до середины мочеточников - 2 слоя: внутренний и наружный. С середины мочеточников и в пузыре - 3 слоя: внутренний, средний, наружный. Наружная оболочка - является адвентициальной, т.е образована соединительной тканью. Мочеиспускательный канал: Предстательная часть: Слизистая оболочка - переходный эпителий. Собственная пластинка: мелкие слизистые железы, сеть венозных сосудов, много эластических волокон. Подслизистая основа содержит сеть широких венозных сосудов. Мышечная оболочка - 2 слоя гладких миоцитов: продольный и циркулярный. Перепончатая часть: Слизистая оболочка - многорядный призматический эпителий; много бокаловидных клеток. Собственная пластинка: мелкие слизистые железы, сеть венозных сосудов, много эластических волокон. Подслизистая основа - содержит сеть широких венозных сосудов. Мышечная оболочка - одиночные пучки гладких миоцитов. Губчатая часть Слизистая оболочка - многорядный эпителий, который в головке члена переходит в многослойный плоский полуороговевающий эпителий. Собственная пластинка: мелкие слизистые железы, сеть венозных сосудов, много эластических волокон. Подслизистая основа - содержит сеть широких венозных сосудов. Мышечная оболочка - одиночные пучки гладких миоцитов. XVIII. ПОЛОВАЯ СИСТЕМА 1. Семенник (яичко). Строение, функции. Яичко снаружи покрыто брюшиной, под брюшинной оболочкой находится капсула из плотной неоформленной волокнистой соединительной ткани – белочная оболочка. На боковой поверхности белочная оболочка утолщается – средостение яичка. От средостения радиально отходят соединительнотканные перегородки, делящие орган на дольки. В каждой дольке находятся 1-4 извитых семенных канальцев, которые в средостении сливаясь между собой продолжаются в прямые канальцы и канальцы сети семенника. Извитой семенной каналец изнутри выстилается эпителиосперматогенным слоем, снаружи покрыт собственной оболочкой. Эпителиосперматогенный слой извитых семенных канальцев состоит из 2-х клеточных дифферонов: спрематогенные клетки и поддерживающие клетки. Сперматогенные клетки – половые клетки на самых разных стадиях сперматогенеза: Второй дифферон эпителиосперматогенного слоя – поддерживающие клетки (синонимы: сустентоциты, клетки Сертоли): крупные клетки пирамидной формы, цитоплазма оксифильная, ядро неправильной формы, в цитоплазме имеются трофические включения и практически все органоиды общего назначения. Цитолемма клеток Сертоли образует бухтообразные впячивания, куда погружаются созревающие половые клетки. Функции: трофика, питание половых клеток; участие в выработке жидкой части спермы; входят в состав гемато-тестикулярного барьера; опорно-механическая функция для половых клеток; под воздействием фоллитропина (ФСГ) аденогипофиза синтезируют андрогенсвязывающий белок (АСБ) для создания необходимой концентрации тестостерона в извитых семенных канальцах; синтез эстрогенов (путем ароматизации тестостерона); фагоцитоз дегенерирующих половых клеток. Придаток яичка (эпидедимис). В придаток яичка семенная жидкость поступает по выносящим канальцам, образующим головку эпидедимиса. Выносящие канальцы в теле органа сливаясь между собой продолжаются в канал придатка. Выносящие канальцы выстланы своеобразным эпителием, где кубический железистый эпителий чередуется призматическим мерцательным, поэтому контур просвета этих канальцев в поперечном срезе складчатый или “зубчатый”. Средняя оболочка выносящих канальцев состоит из тонкой прослойки миоцитов, наружная оболочка – из рыхлой соединительной ткани. Канал придатка выстлан 2-х рядным мерцательным эпителием, потому просвет канала на срезе имеет ровную поверхность; в средней оболочке по сравнению с выносящими канальцами увеличивается количество миоцитов. 2. Семявыносящие пути. Придаток яичка. Семявыносящий проток. Семяизвергательный канал. Семенные пузырьки. Бульбо-уретральные железы. Предстательная железа. Их строение и функции. Возрастные изменения. Семявыносящие пути. К ним относятся прямые канальцы, канальцы сети яичка, выносящие канальцы, канал придатка, семявыносящий и семявыбрасывающий протоки, мочеиспускательный канал. Они обеспечивают депонирование, питание и созревание спермиев, обеспечение их выброса во время эякуляции, эндокринную и секреторную функции. Это трубчатые органы, построенные по общему плану и состоящие из слизистой, мышечной и адвентициальной оболочек. Эпителий слизистой в разных отделах семявыносящих путей различен. В прямых канальцах – однослойный кубический с наличием на апикальных концах микроворсинок и ресничек, в сети семенника – однослойный плоский. В собственной пластинке находятся единичные гладкие миоциты. Выносящие канальцы имеют хорошо выраженные три оболочки: слизистую, мышечную и адвентициальную. В эпителииразличают два типа клеток – реснитчатые и секреторные, имеющие разную высоту, в результате чего просвет канальца неровный. Мышечная оболочка образована несколькими слоями гладких мышц. Канал придатка, образующий его тело, выстлан однослойным двухрядным призматическим эпителием, одни клетки которого на апикальных концах содержат стереоцилии, другие служат камбием (базальные клетки). Средняя и наружная оболочки образованы, соответственно, гладкой мышечной и рыхлой соединительной тканью. Эпителий канала придатка вырабатывает секрет, разжижающий сперму, образует гликокаликс, который покрывает спермии. Удаление гликокаликса при эякуляции приводит к активации сперматозоидов (капацитация). Придаток выполняет также функцию резервуара для накопления спермы. Семявыводящий проток, сменяющий канал придатка, имеет длину около 45 см, главной функцией которого является проведение спермы. Выстлан внутри однослойным двухрядным эпителием с наличием реснитчатых и вставочных клеток. Мышечная оболочка состоит из внутреннего и наружного продольного и среднего циркулярного слоев. Семяизвергающий каналотличается от семявыносящего протока лишь менее развитой мышечной оболочкой. Добавочные железы. К этим органам относятся семенные пузырьки, простата, бульбоуретральные железы. Они вырабатывают секрет щелочного характера, который разжижает спермии и выполняет трофическую роль, а также и эндокринную функцию. Семенные пузырьки. Это парные образования, которые образуются путем выпячивания стенки семявыносящего протока. Их слизистая оболочкавыстлана многорядным призматическим эпителием и образует многочисленные складки. В собственной пластинке, отличающейся обилием эластическим волокон, располагаются концевые отделы слизистых альвеолярных желез. Мышечнаяоболочка образована внутренним циркулярным и внешним продольным слоями гладких мышечных клеток. Наружная оболочка – адвентициальная, представлена рыхлой неоформленной соединительной тканью с обилием эластических волокон. Секрет семенных пузырьков имеет щелочной характер, богат фруктозой, разжижает сперму. Предстательная железа (простата). Это мышечно-железистый орган, массой до 20 гр., в виде муфты, охватывающий семявыводящие пути на уровне впадения семяизвергательного протока в уретру. 3. Яичник. Общая характеристика строения. Особенности строения коркового и мозгового вещества. Овариальный цикл и его гормональная регуляция. Возрастные изменения яичника. Яичники (парный орган) выполняют генеративную (образование женских половых клеток) и эндокринную (выработка половых гормонов) функции. Развитие. Дифференцировка яичника наступает к 6-й неделе эмбриогенеза. В эмбриогенезе яичников усиленное развитие мезенхимы происходит в основании тел первичных почек, при этом редуцируются свободные концы половых шнуров и почечные канальцы, а мезонефральные протоки атрофируются, тогда как парамезонефральные протоки (мюллеровы) становятся маточными трубами, концы которых расширяются в воронки, охватывающие яичники. Нижние части парамезонефральных протоков, сливаясь, дают начало матке и влагалищу. Корковое вещество формируется при врастании половых шнуров от поверхности эпителия полового валика. Мозговое вещество развивается из первичной почки (разрастающейся мезенхимы и кровеносных сосудов мезовария). Строение. С поверхности орган окружен белочной оболочкой, образованной плотной волокнистой соединительной тканью, покрытой мезотелием. Под белочной оболочкой располагается корковое вещество, а глубже — мозговое вещество. Корковое вещество образовано фолликулами различной степени зрелости, расположенными в соединительнотканной строме. Примордиальные фолликулы состоят из овоцита в диплотене профазы мейоза, окруженного одним слоем плоских клеток фолликулярного эпителия и базальной мембраной. Вокруг цитолеммы появляется вторичная, блестящая зона, снаружи от которой располагаются в 1—2 слоя кубические фолликулярные клетки на базальной мембране. Фолликулы, состоящие из растущего овоцита, формирующейся блестящей зоны и слоя кубического фолликулярного эпителия, называются первичными фолликулами. По мере увеличения растущего фолликула окружающая его соединительная ткань уплотняется, давая начало внешней оболочке фолликула. 4. Маточные трубы, матка, влагалище. Циклические изменения органов женского генитального тракта и их гормональная регуляция. Возрастные изменения. Яйцеводы. Маточные трубы или яйцеводы — парные органы, по которым яйцо из яичников проходит в матку. Развитие. Маточные трубы развиваются из верхней части парамезонеф-ральных протоков. Строение. Стенка яйцевода имеет три оболочки: слизистую, мышечную и серозную. Слизистая оболочка собрана в крупные разветвленные продольные складки. Она покрыта однослойным призматическим эпителием, который состоит из двух видов клеток — реснитчатых и железистых, секретирующих слизь. Собственная пластинка слизистой оболочки представлены рыхлой волокнистой соединительной тканью. Мышечная оболочка, следующая за слизистой, состоит из внутреннего циркулярного или спирального слоя и наружного продольного. Снаружи яйцеводы покрыты серозной оболочкой Матка — мышечный орган, предназначенный для осуществления внутриутробного развития плода. Развитие. Матка и влагалище развиваются у зародыша из дистального отдела левого и правого парамезонефральных протоков в месте их слияния. В связи с этим вначале тело матки характеризуется некоторой двурогостью, но к 4-му месяцу внутриутробного развития слияние заканчивается и матка приобретает грушевидную форму. Строение. Стенка матки состоит из трех оболочек: слизистой (эндометрий), мышечной (миометрий) и серозной (периметрии). В эндометрии различают два слоя — базальный и функциональный. Миометрий — состоит из трех слоев гладких мышечных клеток — внутреннего подслизистого, среднего сосудистого с ко-сопродольным расположением миоцитов, богатого сосудами, и наружного надсосудистого. Периметрий покрывает большую часть поверхности матки. В формировании периметрия принимают участие мезотелий, лежащий на поверхности органа, и рыхлая соединительная волокнистая ткань, составляющие прослойку, примыкающую к мышечной оболочке матки. Влагалище. Стенка влагалища состоит из слизистой, мышечной и адвентициалъной оболочек. В составе слизистой оболочки имеется многослойный плоский эпителий, в котором различают три слоя: базальный, промежуточный и поверхностный, или функциональный. Железы в стенке влагалища отсутствуют. Подслизистая основа во влагалище не выражена и собственная пластинка слизистой оболочки непосредственно переходит в прослойки соединительной ткани в мышечной оболочке, которая в основном состоит из продольно идущих пучков гладких мышечных клеток. Адвентициальная оболочка влагалища состоит из рыхлой волокнистой неоформленной соединительной ткани, связывающей влагалище с соседними органами. В этой оболочке располагается венозное сплетение. Циклические изменения влагалища. С началом пролиферации эндометрия, т.е. в постменструальном периоде, во влагалище заметно набухают эпителиальные клетки. На 7—8-й день в этом эпителии дифференцируется промежуточный слой уплотненных клеток, а к 12—14-му дню цикла (к концу постменструального периода) в базальном слое эпителия клетки сильно набухают и увеличиваются в объеме. В верхнем (функциональном) слое влагалищного эпителия клетки разрыхляются и в них накапливаются глыбки кератогиалина. Однако процесс кератинизации не доходит до полного ороговения. В предменструальном периоде деформированные уплотненные клетки функционального слоя влагалищного эпителия продолжают отторгаться, а клетки базального слоя уплотняются. Возрастные изменения. Матка. К концу детородного периода и в связи с приближением климакса, когда гормонообразовательная деятельность яичников ослабевает, в матке начинаются инволютивные изменения, прежде всего в эндометрии. Яичники. В первые годы жизни размеры яичников у девочки увеличиваются преимущественно за счет роста мозговой части. Атрезия фолликулов, прогрессирующая в детском возрасте, сопровождается разрастанием соединительной ткани, а после 30 лет разрастание соединительной ткани захватывает и корковое вещество яичника. Влагалище. После наступления климактерического периода влагалище претерпевает атрофические изменения, его просвет суживается, складки слизистой оболочки сглаживаются, количество влагалищной слизи уменьшается. 5. Молочные железы. Развитие, строение. Особенности желез в период лактации. Эндокринная регуляция желез. Молочные железы по своему происхождению представляют видоизмененные кожные потовые железы. Но по своему функциональному значению они относятся к женской репродуктивной системе, так как обеспечивают первоначальное питание ребенка немедленно после рождения, когда прекращается получение им питательных веществ через плацентарную кровь. Строение. У половозрелой женщины каждая молочная железа состоит из 15—20 отдельных железок, разделенных прослойками рыхлой соединительной и жировой ткани. Эти железы по своему строению являются сложными альвеолярными, и их выводные протоки открываются на вершине соска. Выводные протоки переходят в расширенные молочные синусы, служащие резервуарами, в которых накапливается молоко, продуцируемое в альвеолах. В молочные синусы впадают многочисленные ветвящиеся и анастомозирующие молочные протоки, заканчивающиеся до наступления периода лактации тонкими слепыми трубочками — альвеолярными молочными ходами, которые во время беременности и лактации дают начало многочисленным альвеолам. Молочные синусы открываются на верхушке соска, представляющего собой утолщение кожи. Его эпидермис сильно пигментирован, в базальную часть эпителиального слоя вдаются длинные и часто разветвляющиеся сосочки дермы, в которых располагаются многочисленные инкапсулированные нервные окончания. Обилие этих чувствительных рецепторных окончаний связано с тем, что выделение молока обусловлено рефлексом на раздражение этих окончаний актом сосания. Выработка молока происходит в альвеолах, имеющих вид округлых или слегка вытянутых пузырьков. Железистые клетки альвеол (лактоциты) соеднняются с помощью замыкающих пластинок и десмосом, располагаются в один слой на базальной мембране. По окончании периода лактации молочная железа претерпевает инволюцию, однако часть альвеол, образовавшихся во время предыдущей беременности, сохраняется. Изменения молочных желез половозрелых женщин в течение овариально- менструального цикла. Разрастание концевых отделов отмечается за несколько дней перед овуляцией и продолжается до 20-го дня: с 22—23-го дня пролиферативные процессы прекращаются и альвеолы претерпевают обратное развитие до первых дней менструальной фазы. С 9—10-го дня вновь начинается разрастание ацинусов, но в их клетках нет признаков секреции. Возрастные изменения. У девочек в период полового созревания начинается интенсивное развитие молочных желез. Из разветвленных железистых трубок дифференцируются секреторные отделы — альвеолы, или ацинусы. В течение полового цикла секреторная активность повышается в период овуляции и снижается во время менструации. После прекращения образования овариальных гормонов с наступлением климактерического периода или после кастрации молочная железа претерпевает инволюцию. Регуляция функции молочных желез. Регуляция деятельности функционирующей молочной железы осуществляется двумя основными гормонами — аденогипофизарным пролактином (лактотропный гормон), который стимулирует железистые клетки альвеол (лактоциты) к биосинтезу молока, накопляющегося сначала в млечных ходах и выбрасываемого оттуда во время лактации под влиянием окситоцина — гипоталамического нонапептидного нейрогормона.