IV семестр «Функция и эволюция» БЛОК 1 «Эволюция» – 4 занятия
реклама

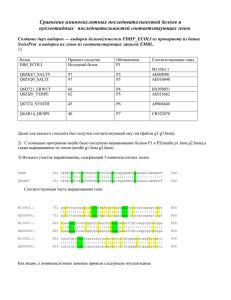
IV семестр «Функция и эволюция» БЛОК 1 «Эволюция» – 4 занятия • • • Молекулярная филогенетика. Задачи и подходы. Лекциясеминар, 13.02. (АБР) Реконструкция филогенетических деревьев. 2 практических занятия, 20.02 и 27.02 (САС, АБР) «Алгоритмы реконструкции филогенетических деревьев». Лекция, семинар, обсуждение полученных результатов, 6.03 (САС). Срок сдачи основного отчета – 5.03 БЛОК 2 «Функции генов и их продуктов» – 4 занятия БЛОК 3 «Эволюция белкового семейства» – 5 занятий Biologists must constantly keep in mind that what they see was not designed, but rather evolved. It might be thought, therefore, that evolutionary arguments would play a large part in guiding biological research, but this is far from the case. Francis Crick What Mad Pursuit (1988) pp.138-139 Вы это уже делали! Прокомментируйте… MYG_HETPO/ MYG_GALGA/ MYG_ALLMI/ MYG_CYPCA/ HBA_HETPO/ HBA1_TORMA HBA_SQUAC/ HBA_LEPPA/ HBA1_BOSMU HBAT_HORSE HBA1_IGUIG HBAZ_CAPHI HBA3_PLEWA HBAD_PASMO HBAD_LIOMI HBA1_XENBO HBA1_PLEWA HBA_CATCL/ HBB1_XENBO HBB_RANCA/ HBB2_XENLA HBB0_MOUSE HBBN_AMMLE HBB_ALLMI/ HBB1_CYGMA HBB_LEPPA/ HBB_SQUAC/ HBB_HETPO/ HBAM_RANCA : : : : : : : : : : : : : : : : : : : : : : : : : : : : : * 20 * 40 * 60 * 80 EWEHVNKVWAVVEPDIPAVGLAILLRLFKEHKETKDLFPKF-KE-IPVQQLGNNEDLRKHGVTVLRALGNILKQ------KGKHSTNVK DWDKVNSVWSAMEANITAVGQNILLRLFEQYPESQSYFPKL-KN-KSLGELKDTADIKAQADTVLKALGNIVKK------KGNHSQPVK EWKHVLDIWTKVESKLPEHGHEVIIRLLQEHPETQERFEKF-KHMKTADEMKSSEKMKQHGNTVFTALGNILKQ------KGNHAEVLK DAELVLKCWGGVEADFEGTGGEVLTRLFKQHPETQKLFPKF-VG-IASNELAGNAAVKAHGATVLKKLGELLKA------RGDHAAILK DRAELAALSKVLAQNAEAFGAEALARMFTVYAATKSYFKDY-KDFT-----AAAPSIKAHGAKVVTALAKACDHL-----D-DLKTHLH NKKAIKNLLQKIHSQTEVLGAEALARLFECHPQTKSYFPKF-SGFS-----ANDKRVKHHGALVLKALVDTNKHL-----D-DLPHHLN DKTAIKHLTGSLRTNAEAWGAESLARMFATTPSTKTYFSKF-TDFS-----ANGKRVKAHGGKVLNAVADATDHL-----D-NVAGHLD DEVLIKEAWGLL-HQIPNAGGEALARMFSCYPGTKSYFPHFGHDFS-----ANNEKVKHHGKKVVDAIGQGVQHL-----H-DLSSCLH DKGNVKAAWGKVGGHAAEYGAEALERMFLSFPTTKTYFPHF--DLS-----QGSAQVKGHGAKVAAALTKAVEHL-----D-DLPGALS DRATVRALWKKMGSNVGVYATEALERMFLGFPSTTTYFLHL--DLS-----LGSTQVKAHGQKVADALTLAVEHL-----E-DLPRALS DKNHIRAIWGHVDNNPEAFGVEALTRLFLAYPATKTYFAHF--DLN-----PGSAQIKAHGKKVVDALTQAVNNL-----D-DIPDALA ERTIILSLWSKISTQADVIGTETLERLFSCYPQAKTYFPHF--DLH-----SGSAQLRAHGSKVVAAVGDAVKSI-----D-NVTSALS EKALVVGLCGKISGHCDALGGEALDRLFASFGQTRTYFSHF--DLS-----PGSADVKRHGGKVLSAIGEAAKHI-----D-SMDQALS DKKLIQQIWGKLGGAEEEIGADALWRMFHSYPSTKTYFPHF--DLS-----QGSDQIRGHGKKVVAALSNAIKNL-----D-NLSQALS DRRLLQASVGKLGCRLEDIGADALNRLLITFPQSKTYFSHF--NLS-----PGSKDIIHQGEKVGKALDSALKHL-----D-DIRGTLS DKKHIKAIMPSIAAHGDKFGGEALYRMFLVNPKTKTYFPTF--DFH-----HNSKQISAHGKKVVDALNEASNHL-----D-NIAGSLS DKHNVKAIWDHVKGHEEAIGAEALYRMFCCMPTTRIYFPAK--DLS-----ERSSYLHSHGKKVVGALTNAVAHI-----D-DIDTAFS DKADVKIAWAKISPRADEIGAEALGRMLTVYPQTKTYFAHW-ADLS-----PGSGPVKHGKKVIMGAIGDAVTKF-----D-DLLGGLA DRQLINSTWGKV--CAKTIGKEALGRLLWTYPWTQRYFSSF-GNLNSADAVFHNEAVAAHGEKVVTSIGEAIKHM-----D-DIKGYYA GGSDVSAFLAKV--DKRAVGGEALARLLIVYPWTQRYFSTF-GNLGSADAISHNSKVLAHGQRVLDSIEEGLKHP-----Z-BLKAYYA EKAAITSVWQKV--NVEHDGHDALGRLLIVYPWTQRYFSNF-GNLSNSAAVAGNAKVQAHGKKVLSAVGNAISHI-----D-SVKSSLQ EKAAITSIWDKV--DLEKVGGETLGRLLIVYPWTQRFFDKF-GNLSSAQAIMGNPRIKAHGKKVLTSLGLAVKNM-----D-NLKETFA BKALITGFWSKV--KVBZVGAZALGRLLVVYPWTZRFFZHF-GBLSSABAVMBBAKVKAHGKKVLBSFSBGLKHL-----B-BLKGAFA ERKFIVDLWAKV--DVAQCGADALSRMLIVYPWKRRYFEHF-GKMCNAHDILHNSKVQEHGKKVLASFGEAVKHL-----D-NIKGHFA ELTIINDIFSHL--DYDDIGPKALSRCLIVYPWTQRHFSGF-GNLYNAEAIIGNANVAAHGIKVLHGLDRGLKNM-----D-NIVDAYA EKQYIVSVFSKI--DVDHVGANTLERVLIVFPWTKRYFNSF-GDLSSPGAIKHNNKVSAHGRKVLAAIIECTRHF-----G-NIKGHLA EKALVNAVWTKT--DHQAVVAKALERLFVVYPWTKTYFVKFNGKFH-----ASDSTVQTHAGKVVSALTVAYNHI-----D-DVKPHFV ELHEITTTWKSI--DKHSLGAKALARMFIVYPWTTRYFGNL-KEFT-----ACSYGVKEHAKKVTGALGVAVTHL-----G-DVKSQFT EKSAVASLWEKIAPQTNKLGAESMERLFKNHPETKSFFSRF--DIS-----PGSQDLLTHGGKIFGALGEAIKSL----------DNLQ 6 g 6 R p F 6 hg 6 : : : : : : : : : : : : : : : : : : : : : : : : : : : : : 81 81 82 81 77 77 77 77 76 76 76 76 76 76 76 76 76 77 80 80 80 80 80 80 80 80 76 75 72 Весна, 2008, А.Б.Рахманинова Молекулярная эволюция (молекулярная филогенетика) Весна, 2008, А.Б.Рахманинова Молекулярная филогенетика ─ изучение филогенеза и эволюции путем анализа нуклеотидных и аминокислотных последовательностей Основные этапы биоинформатического анализа молекулярной эволюции Выбор последовательностей и их выравнивание Построение/выбор эволюционной модели Реконструкция эволюции реконструкция филогенетического дерева оценка силы давления и направления отбора сравнение скоростей эволюции ... Оценка статистической значимости реконструкции ? Весна, 2008, А.Б.Раманинова Что будет? Выбор последовательностей и их выравнивание Построение/выбор эволюционной модели Реконструкция эволюции реконструкция филогенетического дерева оценка силы давления и направления отбора сравнение скоростей эволюции ... Оценка статистической значимости реконструкции ! Весна, 2008, А.Б.Рахманинова Выбор последовательностей и их выравнивание P04443|HBB : MVHFTAEEKAAITSIWDKVDLEKVGGETLGRLLIVYPWTQRFFDKFGNLSSAQAIMGNPRIKAHGKKVLTSLGLAVKNMDNLKETFAHLSE : P07432|HBB : -MGLTAHDRQLINSTWGKVCAKTIGKEALGRLLWTYPWTQRYFSSFGNLNSADAVFHNEAVAAHGEKVVTSIGEAIKHMDDIKGYYAQLSK : : :**.:: *.* *.** :.:* *:***** .******:*..****.**:*:: * : ***:**:**:* *:*:**::* :*:**: 100 * 120 * 140 P04443|HBB : LHCDKLHADPENFKLLGNMLVIVLSSYFGKEFTAEAQAAWQKLVVGVATALSHKYH : 147 P07432|HBB : YHSETLHVDPCNFKRFGGCLSISLARQFHEEYTPELHAAYEHLFDAIADALGKGYH : 146 *.:.**.** *** :*. * * *: * :*:*.* :**:::*. .:* **.: ** gene1(P04443) gene2(P07432) TACCCATGGACTCAGAGG TATCCCTGGACCCAAAGA ** ** ***** ** ** Весна, 2007, А.Б.Рахманинова Выравнивание ? # Length: 502 # Identity: 286/502 (57.0%) # Similarity: 286/502 (57.0%) # Gaps: 119/502 (23.7%) # Score: 678.5 #======================================= CAA32220| AAA49655| CAA32220| AAA49655| CAA32220| AAA49655| CAA32220| AAA49655| CAA32220| AAA49655| 1 atggttcatttcacagctgaggagaaggcagct-atcacaagcatctggg |||| .|||.|||||..|.|| ..|.||||| ||||..||||.||||| 1 atgg---gtttgacagcacatga-tcgtcagctgatcaacagcacctggg 49 50 ataaagt--------ggac--ttggaaaaagttggaggagaaactctggg ..||||| .||| |.|||||| |||| |.||.|| 47 gcaaagtatgtgccaagactataggaaaa-----gagg-----cccttgg 89 90 aaggctcctgattg--tttacccatggactcagaggttctttgacaaa-|.|.||.||| || .|||.||.|||||.||.||.|.|||| | 87 acgtctgctg--tggacttatccctggacccaaagatacttt-----agt 135 136 ---tttggaaacctctcttctgcccaagccatcatgggtaatcccagaat |||||.||||||.....|||..|.|||.|| |.|||.||| 130 tcttttgggaacctcaacagtgctgacgccgtc--------ttccacaat 182 183 caa--------agcccatggcaagaaagtgctgacatccctgggcttggc .|| .||.|||||..|.||.|||.|||||||..|.||...||| 172 gaagcagtggctgctcatggtgaaaaggtggtgacatctattggagaggc 224 46 86 129 171 221 Весна, 2008, А.Б.Рахманинова Выравнивание ? # Length: 502 # Identity: 286/502 (57.0%) # Similarity: 286/502 (57.0%) # Gaps: 119/502 (23.7%) # Score: 678.5 #======================================= # Length: 444 # Identity: 248/444 (55.9%) CAA32220| 1 atggttcatttcacagctgaggagaaggcagct-atcacaagcatctggg 49 # Similarity: (55.9%) |||| 248/444 .|||.|||||..|.|| ..|.||||| ||||..||||.||||| # Gaps: 1 atgg---gtttgacagcacatga-tcgtcagctgatcaacagcacctggg 3/444 ( 0.7%) AAA49655| 46 # Score: 468.0 CAA32220| 50 ataaagt--------ggac--ttggaaaaagttggaggagaaactctggg 89 #======================================= ..||||| .||| |.|||||| |||| |.||.|| AAA49655| 47 gcaaagtatgtgccaagactataggaaaa-----gagg-----cccttgg 86 CAA32220| 1 atggttcatttcacagctgaggagaaggcagctatcacaagcatctggga .|...|||.|||||..|.||..........||||..||||.|||||. AAA49655| 1 ---atgggtttgacagcacatgatcgtcagctgatcaacagcacctgggg CAA32220| 90 aaggctcctgattg--tttacccatggactcagaggttctttgacaaa-135 |.|.||.||| || .|||.||.|||||.||.||.|.|||| | CAA32220| 51 taaagtggacttggaaaaagttggaggagaaactctgggaaggctcctga AAA49655| 87 acgtctgctg--tggacttatccctggacccaaagatacttt-----agt 129 .|||||........|.|...|.|||..|||..|.||.|||.|.||.|||. AAA49655| 48 caaagtatgtgccaagactataggaaaagaggcccttggacgtctgctgt CAA32220| 136 ---tttggaaacctctcttctgcccaagccatcatgggtaatcccagaat 182 |||||.||||||.....|||..|.|||.|| |.|||.||| CAA32220| 101 ttgtttacccatggactcagaggttctttgacaaatttggaaacctctct AAA49655| 130 tcttttgggaacctcaacagtgctgacgccgtc--------ttccacaat 171 ....|||.||.|||||.||.||.|.||||......|||||.||||||... AAA49655| 98 ggacttatccctggacccaaagatactttagttcttttgggaacctcaac CAA32220| 183 caa--------agcccatggcaagaaagtgctgacatccctgggcttggc 224 .|| .||.|||||..|.||.|||.|||||||..|.||...||| CAA32220| 151 tctgcccaagccatcatgggtaatcccagaatcaaagcccatggcaagaa AAA49655| 172 gaagcagtggctgctcatggtgaaaaggtggtgacatctattggagaggc 221 ..|||..|.|||.||.|....|||.....|.|....||.|||||..|.|| AAA49655| 148 agtgctgacgccgtcttccacaatgaagcagtggctgctcatggtgaaaa CAA32220| AAA49655| 50 47 100 97 150 147 200 197 201 agtgctgacatccctgggcttggcagttaagaacatggacaacctcaagg 250 .|||.|||||||..|.||...|||..|.|||.|||||||..||.|.|||| 198 ggtggtgacatctattggagaggccatcaagcacatggatgacattaagg 247 Весна, 2007, А.Б.Рахманинова Выбор последовательностей и их выравнивание P04443|HBB : MVHFTAEEKAAITSIWDKVDLEKVGGETLGRLLIVYPWTQRFFDKFGNLSSAQAIMGNPRIKAHGKKVLTSLGLAVKNMDNLKETFAHLSE : P07432|HBB : -MGLTAHDRQLINSTWGKVCAKTIGKEALGRLLWTYPWTQRYFSSFGNLNSADAVFHNEAVAAHGEKVVTSIGEAIKHMDDIKGYYAQLSK : : :**.:: *.* *.** :.:* *:***** .******:*..****.**:*:: * : ***:**:**:* *:*:**::* :*:**: 100 * 120 * 140 P04443|HBB : LHCDKLHADPENFKLLGNMLVIVLSSYFGKEFTAEAQAAWQKLVVGVATALSHKYH : 147 P07432|HBB : YHSETLHVDPCNFKRFGGCLSISLARQFHEEYTPELHAAYEHLFDAIADALGKGYH : 146 *.:.**.** *** :*. * * *: * :*:*.* :**:::*. .:* **.: ** X14061.1|C : M32456.1|A : ATGGTTCATTTCACAGCTGAGGAGAAGGCAGCT-ATCACAAGCATCTGGG ---ATGGGTTTGACAGCACATGATC-GTCAGCTGATCAACAGCACCTGGG Весна, 2008, А.Б.Рахманинова Что делать? ! tranalign PAL2NAL (http://coot.embl.de/pal2nal/) Весна, 2008, А.Б.Рахманинова Эволюционная модель - математическая модель, описывающей изменения последовательностей во времени. Нужна поправка Число эволюционных событий оценивают, сравнивая 2 родственные последовательности Наблюдаемое число различий между последовательностями меньше реального числа из-за повторных мутаций Резюме: чтобы оценить реальное число событий нужна модель. Однопараметрическая модель Джукса-Кантора (1969) PA(t=0) =PA(0) =1 P A(1) =1-3 C T G A PA(2) = PA(1) (1-3) +(1- PA(1) ) …………….. Djc = -b ln (1 - D/b) Djc – расстояние по Джуксу-Кантору (среднее ожидаемое число замен за время t) b – константа, для нуклеотидных последовательностей b=3/4 D – неоткорректированные расстояния (в простейшем случае, это доля несовпадающих букв) Однопараметрическая модель Джукса-Кантора Вероятность А в момент t PA(t=0) =PA(0)=1 PA(1) =1-3 PA(2) = PA(1) (1-3) +(1- PA(1) ) PA(t+1) = PA(t) (1-3) + (1- PA(t)) dPA(t) = - 4 P + —— A(t) dt PA(t) = 1/4 + (PA(0) -1/4) e -4t PA(t) = 1/4 + 3/4 e-4t для PA(0)=1 PA(t) = 1/4 - 1/4 e-4t для PA(0)=0 Расстояние до предковой последовательности 1. Модель предполагает, что в 1 позиции в единицу времени происходит 3 замен. 2. Прошло время t, Djc – общее число замен/позицию за время t. 3. Доля совпадений/позицию между предковой и настоящей последовательностью? 4. Доля несовпадений/позицию между предковой и настоящей последовательностью? 5. Расстояние между предковой и настоящей последовательностью в смысле ожидаемого числа замен за время t (Djc ) - ? Двухпараметрическая модель М.Кимура (1980) C T G A distance = -0.5 ln[ (1-2P-Q)*sqrt(1-2Q)] P = transitions/npos Q = transversions/npos трансверсия транзиция Чем точнее модель, тем лучше результат? ? по материалам Simon Whelan, Pietro Lio and Nick Goldman "Molecular phylogenetics: state-of-theart methods for looking into the past" TRENDS in Genetics Vol.17 No.5 May 2001 Сравнение скоростей эволюции разных генов 1. Thr-Ser-Ala… 1.Thr-Ser-Ala… 2. Thr-Ser-Ala… 2.Pro-Arg-Asp… 1. ACT AGT GCC… 2. ACA AGC GCT… ? 1. ACT AGT GCC… 2. CCT AGA GAC… Весна, 2008, А.Б.Рахманинова KA/KS – мера давления естественного отбора Несинонимичные замены Синонимичные замены Сравниваем 2 наблюдения: ATG GGG GCT GGG ATA GGA GAT GGA KA 1 1 Ks 1 1 KA/KS =1 KA/KS =1 => одно и то же ? Необходимо нормировать на «возможности» последовательности: Несинонимичные сайты Синонимичные сайты PA Ps 5 1 4 2 Нормируем на мутабильность последовательностей: KA = KA/PA KS = Ks/Ps KA/KS = 1/5 KA/KS = 1/2 KA/KS – мера давления естественного отбора KA – число несинонимичных замен на 1 несинонимичный сайт Ks – число синонимичных замен на 1 синонимичный сайт Пример: Человек Thr Val Asp Gly …. ACT GTT GAT GGT …. Всего Число несинонимичных сайтов 110 110 11 2/3 110 8 2/3 Число синонимичных сайтов 001 001 00 1/3 001 3 1/3 Шимпанзе Thr Val Glu Gly …. ACA GTC GAA GGT …. Всего Число несинонимичных замен 0 0 1 0 1 Число синонимичных замен 1 1 0 0 2 Ka= 1 / 8.67= 0.12 Ks= 2/ 3.33= 0.6 Ka/Ks=0.12 / 0.6 =0.19 (стабилизирующий отбор?) KA/KS – мера давления естественного отбора KA – число несинонимичных замен на 1 несинонимичный сайт Ks – число синонимичных замен на 1 синонимичный сайт Пример: Человек Thr Val Asp Gly …. ACT GTT GAT GGT …. Всего Число несинонимичных сайтов 110 110 11 2/3 110 8 2/3 Число синонимичных сайтов 001 001 00 1/3 001 3 1/3 Шимпанзе Thr Val Glu Gly …. ACA GTC GAA GGT …. Всего Число несинонимичных замен 0 0 1 0 1 Число синонимичных замен 1 1 0 0 2 Ka= 1 / 8.67= 0.12 Ks= 2/ 3.33= 0.6 Ka/Ks=0.12 / 0.6 =0.19 (стабилизирующий отбор?) KA/KS, опять все не просто.... KA/KS – мера давления естественного отбора KA – число несинонимичных замен на 1 несинонимичный сайт Ks – число синонимичных замен на 1 синонимичный сайт KA / Ks << 1 отрицательный (стабилизирующий) отбор KA / Ks 1 нейтральная эволюция KA / Ks > 1 положительный (движущий) отбор KA/KS; какие бывают? Из Hurst, L.D. (2002) Trends in Genet. 18, 486-487 КАТЕГОРИИ ГЕНОВ ЧЕЛОВЕКА, ОБНАРУЖИВАЮЩИХ ПРИЗНАКИ ПОЛОЖИТЕЛЬНОГО ОТБОРА ВЗАИМОДЕЙСТВИЕ ПАТОГЕН-ХОЗЯИН ВОСПРОИЗВОДСТВО ПРИСПОСОБЛЕНИЕ К ПИТАНИЮ ВНЕШНОСТЬ СЕНСОРНЫЕ СИСТЕМЫ ПОВЕДЕНИЕ ОРГАНИЗАЦИЯ МОЗГА НЕИЗВЕСТНОГО НАЗНАЧЕНИЯ E.J. Vallender, B.T. Lahn. Hum.Mol.Gen. 2004, V.13, Rev.Issue 2, R245-R254 Ждите лекций А.В.Алешина по теории эволюции и лекций М.С.Гельфанда по сравнительной геномике!!