Лекция 3 - DNA punctuation
реклама

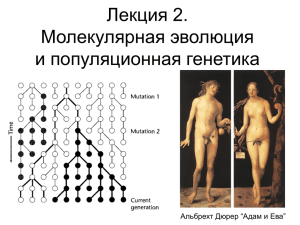
Лекция 3. Генеалогические деревья и коалесценция Альбрехт Дюрер “Адам и Ева” Теория коалесценции • Основана на генеалогических деревьях гена • Рассматривает процесс эволюции назад во времени на основании процессов нейтральности и генетического дрейфа • История любого аллеля (мутации) в популяции начинается с мутации в одном индивидууме • Любая мутация может быть отслежена назад, до того момента, когда она возникла в одном индивидууме, то есть, когда “ДНКпоследовательности” сходятся к одной Теория коалесценции • Можно строить генеалогические деревья, которые будут отражать происхождения различных частей генома • Можно предсказать генеалогическую структуру данного локуса (геномной области), которая эволюционировала нейтрально • Генеалогические структуры, которые отличаются от ожидаемой нейтральной эволюции, свидетельствуют о популяционных демографических событиях (феномен “бутылочного горлышка”, расселение) или действии отбора (селективной преимущество, балансирующий отбор) Теория коалесценции Cвязь с генетическим дрейфом • Генетический дрейф указывает на то, что взаимосвязь размера популяции с ее генетическим разнообразием независима от действия сил отбора. • Если мы знаем “нейтральное ожидание” разнообразия, то мы можем измерять отклонения от него. Ожидание при нейтральности Ожидание при “бутылочном горлышке” Спектр частоты сайтов • число синглтонов, даблтонов, триплтонов в данном наборе последовательностей • обладает ожидаемым распределением (некоторой пропорцией каждого типа полиморфизма) при условии наличия конкретной эволюционной истории данного локуса (геномной области) положительного отбора балансирующего отбора (отбор на поддержание гетерозиготности) нейтральной эволюции демографической истории (бутылочного горлышка и др) Сректр частот сайтов Митохондриальная Ева (Mt MRCA) MRCA - most recent common ancestor • Жила около 200,000 лет назад,возможно, в Восточной Африке • Жила гораздо раньше, чем, по оценкам. произошла миграция из Африки - где-то между 95,000 и 45,000. Y-хромосомный Адам (Y-MRCA) • 60,000 years ago - Map shows first migratory routes taken by humans, based on surveys of different types of the male Y chromosome. "Adam" represents the common ancestor from which all Y chromosomes descended - Research based on DNA testing of 10,000 people from indigenous populations around the world Модель теории коалесценции - N- число индивидуумов в настоящем поколении - Пусть все индивидуумы могут равновероятно иметь одинаковое число потомков. - Вероятность, что любые два индивидуума из настоящего поколения имеют общую мать равна 1/N - Вероятность, что у них разные матери - (1-1/N) Вероятность, что ближайший общий предок 2-х индивидуумов жил T поколений назад: (T-1) поколение - разные матери, а в T-поколении мать общая N - среднее Аналогично, время коалесценции для N индивидуумов - 2N Популяционная генетика • Теория и методы для понимания генетической вариации в популяциях и как эта вариация изменяется под действием отбора, дрейфа и мутаций. • Классическая популяционная генетика – Взгляд вперед: “какова вероятность, фиксируется ли преимущественный аллель, если действует сильный отбор и частота аллеля 70%”? • Теория коалесценции – Взгляд назад: “Имея данные полиморфизма ДНК последовательностей, какова вероятность, что в данной области генома возникла преимущественная мутация, которая впоследствии закрепилась”? Фиксация нейтральных мутаций • • “Думаем назад”: вероятность того, что мутация произошла в гене, который стал общим предком всех присутствующих - 1/N Кимура 1983 вперед” - Теория нейтральной эволюции Теперь “думаем - только мутация в этом одном гене -будет главный результат: фиксирована. Таким образом, процент нейтральных мутаций в настоящем поколении, который будет фиксирован - pfix=1/N. • скорость фиксации нейтральных мутаций равна Пусть u - вероятность возникновения новой мутации в одном сайте ДНК в скорости самих мутаций и не зависит от размера каждом поколении популяции • Nu - среднее число мутаций, возникающее в одном сайте ДНК во всей популяции • Cкорость фиксации нейтральных мутаций - это скорость, с которой мутации возникают, умноженное на вероятность фиксации каждой мутации - ufix = Nu*pfix=u Следующая лекция • Модели эволюции нуклеотидных последовательностей. Модель Джукса-Кантора. Двупараметрическая модель Кимуры. Модель Хасегава-Кишино-Яно. Разброc скоростей эволюции позиций. • Модели эволюции аминокислотных последовательностей. Частоты аминокислотных замен. LOG-ODDs матрицы счета. Матрицы PAM, связь с физикохимическими свойствами аминокислот. Матрицы BLOSUM, JTT. Редактор vi