Адаптация, память и гомеостатическая мощность биосистем определяют наличие и характер их реакций
реклама

Адаптация, память и
гомеостатическая мощность
биосистем определяют наличие
и характер их реакций
Загускин Сергей Львович,
доктор биологических наук,
академик МАЭН,
член Проблемной комиссии по
хронобиологии и хрономедицине РАМН,
член лазерной ассоциации
Лаборатория хронобиологии НИИ физики
Ростовского государственного университета
344022, Ростов-на-Дону, а/я 3408, т.(8632)-227765,
факс (8632)-434044, E-mail: zag@ip.rsu.ru
«Во всем царит гармонии закон, и в мире все суть ритм,
аккорд и тон»
Дж. Драйден
«В числе основных законов мира необходимо поставить
закон периодичности или ритма»
В.М.Бехтерев
«Основные проблемы патологии - это проблемы живой клетки»
«Понимание динамики протоплазмы могло бы объяснить
действие различных физических и химических факторов»
«Сама природа ответного механизма требует, чтобы
блокировать чрезмерно сильную реакцию и обращение
коллоидных изменений»
« Кортикальный слой восстанавливает утерянный им кальций,
а протоплазма внутри клетки вновь приходит в более
жидкое состояние»
«Возможно ли при помощи циклических коллоидных изменений
понять ритмические процессы, происходящие, например,
при сердечных сокращениях?»
Л. Гейльбрун, 1957г.
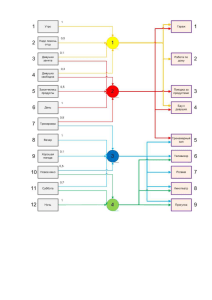
РИТМЫ ЗОЛЬ-ГЕЛЬ ПЕРЕХОДОВ В КЛЕТКЕ КАК
АКЦЕПТОРЫ И РЕЦЕПТОРЫ ВНЕШНИХ ВОЗДЕЙСТВИЙ
Физические воздействия
?
Первичные
акцепторы
Энергетический
обмен
клетки
Са депо
t°C, рН, Росм
Н2О
Гель
Кровоток
Химические
воздействия
1О
Золь
Саi
Pm
МП
2
Биосинтез
Рецепторы
клеточной
мембраны
ЦН
Функция
Прямые функциональные связи,
Обратные связи,
Энергетические связи
«Внутриклеточная подвижность имеет универсальное
биологическое значение, являясь существенным
инструментом регуляторных процессов»
Г.М. Франк, 1962г.
« Структурная подвижность может быть мощным регулятором
интегративной функции клетки в целом»
Г.М. Франк, 1970г.
« Два зрелища меня неизменно восхищают и волнуют:
танец маленьких лебедей и делящиеся хромосомы»
« Выход один – создавать машины, которые могли бы
вместо человека «смотреть» в микроскоп»
« Пройдет немного времени, и автоматы-анализаторы
живых структур совершат в биологии такой же переворот,
какой около 300 лет назад произвел микроскоп»
« Эпоха машинной биологии не за горами»
Г.М. Франк
Дифференциальная интерферометрия живого
нейрона. Регистрация ритмов агрегации
ретикулюма (золь-гель структур)
Нервная клетка механорецептора речного рака.
Регистрация электрической активности, потребления
кислорода, агрегации митохондрий, биосинтеза белка и
других структурно-химических ритмов
мышцы
рО2
электрод
З
нерв
Тело нейрона
З
ганглий
аксон
70 мкм
Камера на столике микроскопа
З- зажимы
З
Интерферометрия живого нейрона с полным раздвоением
изображения. Регистрация ритмов содержания белка (сухого
веса) в покое клетки и при физических воздействиях
Фотоэлектрический метод:
Ф0-Ф1=А{cos2φ-Lcos2(φ-Δ/2)}+M
Фотографический метод:
(S0-S1)/γ=lg{cos2φ/Lcos2(φ-Δ/2)}-M
Увеличение амплитуды ритмов зольгель переходов при облучении лазером
и стимуляции энергетики клетки
Зонд
А
ВВ
1
2
С
А-Метод микрокиноденситографии
В- агрегаты митохондрий
С- увеличение амплитуды колебаний
агрегации-дезагрегации митохондрий
(оптической плотности 1-2) и уровня
золя относительно геля (разжижения
цитоплазмы) при стимуляции
сукцинатом натрия (или лазером)
энергетики клетки.
Синхронные колебания энергетического обмена
и биосинтеза в клетке
Ядро
Д
Я
Я
Д
А
Околочасовые ритмы
(средний период около 30
мин) белка в разных
зонах клетки: Д - между
дендритами и ядром,
Я - в зоне ядра,
А - между ядром и аксонным
холмиком по поглощению
при 265 нм. Калибровка:
оптическая плотность - 0,1,
время - 30 мин.
Аксон
Аксон
Прижизненная интерферометрия одного и того
же нейрона в фазе снижения содержания белка
околочасового ритма (слева) и в фазе его
увеличения (справа). Видно изменение сдвига
фазы волны света над ядром тела нейрона на
уровне двух ядрышек и отсутствие изменений в
участке отходящего вниз аксона.
Энергетическая зависимость возбудимости нейрона и
пластическое закрепление его адаптации
А
А Частотограмма импульсной активности
нейрона при раздражении с периодом 5
мин.
Видно быстрое снижение возбудимости с
начала раздражения и быстрое триггерное
восстановление возбудимости через 90 мин.
Б
Б Прижизненная ультрафиолетовая
цитоспектрофотометрия нейрона.
Графики колебаний поглощения при длине
волны 265 нм в указанных на схеме участках
тела нейрона.
Видно первичное увеличение поглощения
(увеличение концентрации РНП) в участках
над ядром и постепенная фазовая
синхронизация с ними колебаний
поглощения в других участках клетки в
направлении к аксону.
Фазовый гель-золь переход
с потреблением энергии на границе тела
клетки и аксонного холмика
А
С
А- прижизненная морфометрия нервной клетки
В
(метод дифференциальной интерферометрии),
В- «барьер» митохондрий на границе сомы и
аксонного холмика нейрона при его торможении
С- изменение отношения диаметров сомы и
аксонного холмика-(САИндекса),определяющего
декремент генераторного потенциала и
изменение частоты импульсной активности
Различие при импрегнации серебром морфологии золь-гель структур в
цитоплазме и в ядре нервной клетки, зафиксированной в состоянии покоя
(П), импульсной активности, вызванной постоянным умеренным в
течение 20 мин (В-20мин) или ритмическим сильным раздражением в
течение 2 мин (БР-2мин)
П
В-20мин
БР-2мин
Окраска
на ДНК
по
Фельгену
Рентгеноспектральный (электронно-зондовый) микроанализ
химических элементов в инфузории и цитохимический анализ
магния в теле нейрона и в нервных волокнах речного рака
А- Частотограммы импульсной активности нейрона.
Вызванные колебания имеют период в 6, 10 и более раз
больший, чем ритм раздражения с Т=0,5;1;2;11 и 29с.
В- вызванные медленные ритмы можно подавить
другой частотой раздражения.
С- раздражение с Т=0,5с вызывает колебания
частоты, синхронизацию золь-гель переходов,
колебаний Са цитозоля и возбудимости нейрона
с большей амплитудой, чем раздражение с Т=29с.
Д- Осциллограммы импульсной активности нейрона при
выработке временной связи на подпороговое
электрическое раздражение и подкреплении адекватным
механическим раздражением
А
Д
С
В
Зависимость функциональных и биосинтетических реакций клетки
от фазы энергообеспечения
%
А
А Колебания микрогетерогенности (d ) и
D
d
имп/с
1
2
30
60
90 МИН.
Б
Б Частотограммы импульсной активности
нейрона при разражении с частотой 1/с.
1- после предварительной активации
энергетического обмена умеренным
возбуждением в течение 10 мин.
2- сразу после увеличения исходной частоты
без активации энергетики.
20 с.
имп/с
среднего поглощения (D) при длине волны
265 нм в теле нейрона при возбуждении.
Снижение d означает увеличение золя
относительно геля при уменьшении
агрегации митохондрий и ретикулюма и
сответствует увеличению концентрации РНП.
С
имп/с
60
50
40
30
20
10
0
C Частотограммы импульсной активности
1
2
3
4
5
6
7
Величина раздражения
энергия выше
8
энергия ниже
симметричных нейронов в фазе повышения
энергетического обмена и в фазе снижения
энергообеспечения. Порог реакции ниже, а
блок импульсации наступает при большей
силе раздражения, если энергия выше .
Зависимость направленности реакции клетки (объема,
агрегации ретикулюма и золь-гель структур) от исходного
уровня ее энергетики (фазы биоритма +Е или - Е)
+Е
+Е
-Е
-Е
При исходно высоком
энергетическом обмене (+Е)
увеличивается объем клетки и
уровень золя относительно
геля, агрегация ретикулюма и
концентрация кальция в
цитозоле уменьшаются.
Частотограммы импульсной активности нервной
клетки механорецептора рака при
изолированном воздействии ИК лазером
(пунктир) и сочетании его с адекватным
(сплошная линия) раздражением с периодом 29с.
Появление медленных с периодом Т=
2-3 мин. колебаний частоты импульсов
в ответ на лазерное облучение Т=29с
и сохранение их в последействии.
Сочетание лазерного и адекватного
механического раздражения с тем же
периодом. Каждое механическое
раздражение вызывает увеличение
с тем же периодом частоты на 4-5имп/с.
Снова изолированное лазерное
воздействие вызывает более частые
колебания частоты (и возбудимости) с
меньшей амплитудой.
Изменение периодов колебаний частоты
импульсов отражает изменение ритмов
фазовых золь-гель переходов и ритмов
концентрации кальция в цитозоле с
сохранением в последействии.
Выработка временной связи на одиночном
изолированном нейроне после усиления его
энергетического обмена
1
2
3
4
1- частотограмма импульсной
активности нейрона при действии
исходно подпорогового
электрического (лазерного)
воздействия (пунктирная отметка)
и подкрепляющего механического
раздражения (сплошная отметка),
2- сочетанное воздействие,
3- после прекращения подкрепляющего
механического воздействия видны
ответы на только одно электрическое
раздражение и следовые колебания
средней частоты после прекращения
обоих видов раздражения,
4- ответы нейрона на включение
только ранее подпорогового
электрического воздействия
Физико-космический уровень (надбиосферный)
x5
ФСРБ
ФСОБ
y4
w4
v4
x4
ФСРО
y3
v3
ФСО
x3
w2
Ансамбли
z1
PКК
ОКК
y1
z0
Биоценозы
w3
z2
y2
k4
v2
x2
k3
Организмы
u2
k2
Клетки
w1
v1
x1
p 4 l4
p3
l3
k1
БСРЦ
u0 v 0 w 0
БИОМЫ
r4
Консорции
Популяции
r3
s2
m2
p 2 l2
ОРГАНЫ
ТКАНИ
r2
s1
РМК
m1
u1
ЗОНЫ
s3
m3
u3
Основные уровни:
Биосфера, Биоценозы,
Организмы, Клетки,
БСРЦ-биосинтетические
s4
m4
u4
z3
Семьи,стада
k5? p5?
u5
Биосфера
z4
Уровни эволюционной
интеграции биосистем
p1
l1
ОМК
s0
r0
r0 Гены, вирусы
Физико-химический уровень (предбиологический)
r1
саморедуплицирующиеся
циклы золь-гель структур
Промежуточные функциональные уровни: ФСРБфункциональные системы
разнородных биоценозов,
ФСОБ - функциональные
системы однородных биоценозов, ФСО-функциональные
системы организма, РККразнородные компартменты
клетки, ОКК-однородные
компартменты клетки.
Промежуточные структурные
уровни: РМК-разнородные
микроструктуры клетки,
ОМК -однородные
микроструктуры клетки.
Энергетические потоки
на: x,y,z-функциональные и
p,r,s-структурные изменения
u,v.w-параметрическую
регуляцию функции,
k,l,m-параметрическую
регуляцию структуры.
Основные уровни
биологической
интеграции
Энергетические потоки
Функция
Структура
Периоды колебания или длительность
переходных процессов
Расход Вход Расход Вход
Реликтовые
Основные
Координации
k5 ?
m4
l4
k4
m3
l3
k3
m2
l2
k2
m1
l1
k1
2.3 млрд. лет
240 млн. лет
24 млн. лет
7.4 млрд. л
740 млн. л
75 млн. л
23 млрд. лет?
800 тыс. лет
80 тыс. лет
8 тыс. лет
2.4 млн. л
250 тыс. л
25 тыс. л
7.4 млн. лет
260 лет
26 лет
2 года 8 мес.
800 лет
82 года
8 лет 4 мес.
2.5 тыс. лет
1 мес.
3 дня
8 час.
3 мес.
10 дней
24 час.
10 мес.
15 мин.
1.5 мин.
9 с.
50 мин.
5 мин.
30 с.
150 мин.
300 мс.
30 мс.
3 мс.
1 с.
100 мс.
10 мс.
3 с.
100 мкс.
300 мкс.
1 мс.
x5
z4
y4
x4
z3
y3
x3
z2
y2
x2
z1
y1
x1
u5
w4
v4
u4
w3
v3
u3
w2
v2
u2
w1
v1
u1
w0
v0
z0
u0
p5 ?
s4
r4
p4
s3
r3
p3
s2
r2
p2
s1
r1
p1
s0
r0
До лечения
После лечения
До лечения
После лечения
Больной П., группа К2
Больной К., группа К1
Суточный график ЧСС
Тахограммы
Больной Г., группа О1
SDNN
rMSSD
Суточный спектральный анализ
R-R интервалов ЭКГ
pNN50
Почасовые графики
Различие показателей ЭКГ 5-ти минутных
записей одного и того же пациента З., 63г
17.12.2002г
в 16час.07мин.(слева) и в 16час.50 мин.(справа)
Индекс 73% Вы в прекрасной форме
Индекс 39%- Вам следует обратить
внимание на свое здоровье
До лечения
После лечения
Фрактальная
размерность R-R
интервалов ЭКГ
Индекс Херста
Нормированный
показатель
Фишера
Критерии оптимальности параметров
квантовой (лазерной) терапии
Гармоническая фиксированная
частота лазерного воздействия
Плотность
мощности
Биоритм
энергообеспечения
Знак (+)
реакции (--)
Кривая Лапика-Вейса
Время
Выполняется ли закон Бунзена-Роско?
Оптимальны параметры только
в режиме биоуправления.
Факторный анализ значений
Ес, Т, S, λ не корректен из-за
неаддитивности реакций ответа.
Биорезонанс возможен только
при многочастотной
биоритмологической
биосинхронизации.
Критерии оптимальности
параметров лазерной терапии
Переход части геля в золь
Активность СОД
Уровень микроциркуляции
Продукция Т-лимфоцитов
Нормализация ЧП/ЧД
Фрактальная размерность ЧП
Эффект воздействия лазера
0,3 дж/см2 = Ес мВ/см2 х Тмин
0,5 х10
1х5
5х1
нет
+
++
+
+
нет
нет
++
+
нет
++
+
++
+
нет
++
+
нет
Зависимость знака ответной реакции от
исходного состояния и фазы ритма
энергетики клетки, ткани, органа, организма
Качели энергообеспечения
Ян
Инь
Вдогонку
Фаза увеличения
кровенаполнения
= увеличение
биосинтеза
Навстречу
Фаза снижения
кровенаполнения
= деструкция
Соотношение 1-2-3 глубин амплитудной модуляции
физиотерапевтического воздействия меняют в
зависимости от характера патологии (гипоксия,
артериальная или венозная гиперемия) и наличия
дисбаланса артериальной и венозной частей
капиллярного русла
1.Тремор 7-13Гц
систола сердца
2. Пульс
3. Дыхание
вдох
диастола
выдох
Суммарный
сигнал (1,2,3)
биоуправления
АППАРАТ БИОУПРАВЛЯЕМОЙ КВАНТОВОЙ
ХРОНОТЕРАПИИ «РИКТА-05»
•
•
•
Датчик
пульса
Терминал:
8 красных,
1 ИК лазер,
магнит
Датчик
дыхания
Дисплей:
ЧП, ЧД, ЧП/ЧД,
биотаймер,
1-4 режим
•
•
Хронодиагностика и
контроль состояния
пациента во время
сеанса хронотерапии с
биоуправляением по
отношению частоты
пульса (ЧП) к частоте
дыхания (ЧД).
Норма: 3<ЧП/ЧД <5.
Режим 1-4 : разные
соотношения глубин
модуляции по сигналам пульса,
дыхания, тремора.
С учетом скорости пульсовой
волны и места патологии
вводят задержку по сигналу
пульса.
При воздействии в проекции
миокарда включают реверсию
знака сигнала пульса.
Выводы:
«Привычка свыше нам дана, замена ……
(Биологическим эффектам солнечной активности) она»
А.С. Пушкин
1.Способы снижения чувствительности к
привычным внешним воздействиям, не
имеющим сигнальной биологической
значимости:
А)Морфологические, метаболические и
физиологические способы повышения порогов
и смещения диапазона восприятия, защитного
экранирования,
Б)Противофазные эндогенные колебания
высвобождения и связывания ионов кальция,
закрепленные морфологическими параметрами
внутриклеточных кальциевых депо и
соответствующими ритмами фазовых золь-гель
переходов, направленных на
гомеостатирования концентрации кальция в
цитозоле клетки.
Норма
Гомеостатическая мощность биосистемы N-уровня –
диапазон допустимых отклонений параметров по
амплитуде и длительности при Т функциональных
изменений меньше Т структурного восстановления
Е
Старение, локальная патология
Регуляция по отклонению
Системная хроническая патология,
Регуляция по возмущению
Устойчивость = «работа против
равновесия» (Э. Бауэр) =
«Волчки жизни» иерархии биосистем
Т
Вывод 2
Величина и знак
(направленность)
реакции биосистемы
зависят от фазы ритма
Клетки
энергообеспечения, от
исходного состояния
Ткани
биосистемы при
по
Органы регуляции
возмущению и только
при регуляции по
отклонению реакция
ы
Организм
имеет нормализующий,
корректирующий фазу
ритма характер.
Зависимость реакции клетки, органа, организма
от интенсивности и длительности воздействия
Постоянное
Реакция
Импульсное
1
0,8
0,6
0,4
0,2
0
Биоуправление
Биоуправление
- нет адаптации
Длительность
Вывод 3:
Законы Арндта-Шульца,
Вебера-Фехнера, БунзенаРоско и Лапика-Вейса
выполняются при исходно
(и условно) постоянном
уровне чувствительности
(возбудимости нервных и
мышечных клеток по Н.Е.
Введенскому).
При исходно
колебательном уровне
чувствительности
(возбудимости) знак
и величина ответа
зависят от фазы ритма
энергообеспечения реакции.
4. Повышение чувствительности к биологически значимым
сигнатурным воздействиям происходит за счет обучения
(повторяемости) не к определенным частотам, а к образам многочастотным многомодовым воздействиям с
инвариантным соотношением периодов колебаний,
соответствующим иерархии биоритмов биосистемы.
Увеличение фрактальной размерности обеспечивает восприятие
сверхслабых сигналов- биологических кодов на основе
многочастотного параллельного резонансного захвата.
Не так
страшна
солнечная
активность,
как ее…..