Лекция №3 Механизмы клеточного деления. Митотический цикл и его регуляция. Митоз. Мейоз.
реклама

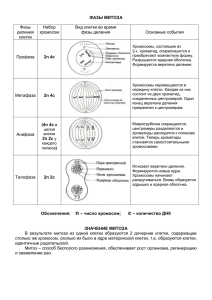
Лекция №3 Механизмы клеточного деления. Митотический цикл и его регуляция. Митоз. Мейоз. Как постулирует клеточная теория, увеличение числа клеток происходит исключительно за счет деления исходной клетки, предварительно удвоившей свой генетический материал. Это – главное событие в жизни клетки как таковой, а именно завершение воспроизведения себе подобного. Деление клетки процесс неслучайный, строго генетически детерминированный. Как известно, клеточные деления бывают двух типов: митоз (кариокинез, непрямое деление) и мейоз. Мейоз используется лишь в одном случае: так происходит последнее деление предшественников половых клеток. Остальные деления осуществляются путем митоза (все предыдущие деления предшественников половых клеток, а также все деления соматических клеток). Прямое деление клеток, или амитоз, достоверно описано только при делении полиплоидных макронуклеусов инфузорий. По способности к делению все клетки взрослого организма подразделяются на 3 типа: 1. Митотические, т.е. постоянно делящиеся клетки (клетки базального слоя эпителия, гемопоэтические клетки начальных стадий созревания, сперматогонии и др. 2. Условно постмитотические клетки. Это неделящиеся клетки, сохранившие способность к делению при действии определенных стимулов. Чаще всего деления возобновляются при регенерации соответствующего органа или ткани (клетки печени, стволовые клетки костных, скелетных мышечных и др. тканей. 3. Постмитотические клетки – неделящиеся клетки, окончательно утратившие способность делиться (клетки всех слоев эпидермиса, кроме базального; нервные клетки; клетки сердечной мышцы; волокна скелетных мышц). Период жизни клетки от одного деления до другого называется митотическим (клеточным) циклом. Жизненный цикл митотически делящейся клетки состоит из четырех стадий: S-период (синтетический): В ядре происходит репликация (удвоение) ДНК и хромосомных белков. Содержание ДНК в ядрах соматических клеток (клеток тела) в 2 раза больше чем в ядрах зрелых половых клеток. Соматические и половые клетки отличаются и по количеству хромосом. Соматические обозначаются как 2n2c, половые nc. В синтетический период количество ДНК постепенно возрастает от 2с до 4с. Число хромосом не изменяется (2n), но каждая содержит 2 сестринские хроматиды. В цитоплазме происходит удвоение центриолей. G2-период (постсинтетический, или премитотический). Не очень продолжительный, включат синтез ряда веществ, необходимых для прохождения митоза. В частности, в это время синтезируется белок микротрубочек тубулин, необходимый для формирования веретена деления. Содержание ДНК в этот период – 4с. Митоз. Включает 4 фазы: профазу, метафазу, анафазу и телофазу. Происходит деление, приводящее к образованию двух диплоидных клеток (2n2c). G1-период (пресинтетический, или постмитотический). В этот период восстанавливается содержание цитоплазматических белков и, как следствие, происходит рост клетки (до размера материнской). Кроме того, происходит синтез белков, необходимых для репликации. Содержание ДНК в клетке – 2n. Именно в этот период принимается решение о вступлении клетки в очередной клеточный цикл или о прекращении делений. Стадии G1, S, G2 вместе составляют интерфазу. Во время митотического цикла смена стадий происходит строго упорядочено. Делящиеся клетки могут продолжать делиться или временно прекратить деления, выйти из цикла. О таких клетках говорят, что они вступили в G0период. Кроме того, в зависимости от ряда обстоятельств, клетка может вступить в процесс дифференцировки, включить механизм самоуничтожения (апоптоз), либо подвергнуться бласттарсформации, т.е. превратиться в опухолевую клетку. Митоз. Митоз – (от греч. Mitos – нить), непрямое деление, основной способ деления эукариотических клеток. Биологический смысл митоза заключается в строго одинаковом распределении редуплицированных хромосом между дочерними клетками, что обеспечиваетобразование генетически равноценных клеток и сохранении преемственности в ряду поколений. У клеток, вступивших в цикл деления, фаза собственно митоза, непрямого деления, занимает относительно короткое время, около 0,1 времени клеточного цикла. Митоз состоит из четырех основных фаз: профаза, метафаза, анафаза, телофаза. Митоз представляет собой непрерывный процесс, поэтому границы между фазами установить достаточно трудно. Профаза. В нее входят клетки из G2-периода интерфазы, они после репликации в S-периоде содержат удвоенное количество ДНК (4с) по сравнению с исходной клеткой в G1-периоде, соответствующее таковому у тетраплоидной клетки. В начале профазы в ядре начинают выявляться тонкие нити – профазные хромосомы. Это результат процесса конденсации хромосом, который совпадает с падением их транскрипционной активности. По мере прохождения профазы хромосомы укорачиваются и утолщаются. Число хромосом соответствует диплоидному количеству, но по содержанию ДНК ядра профазных клеток тетраплоидны. Каждая профазная хромосома состоит из двух взаимноспирализованных хроматид. Сестринские хроматиды связаны друг с другом с помощью белков когезинов. Таким образом, число хроматид (4n) в профазе точно соответствует количеству ДНК (4с). Как следствие конденсации синтез РНК на хромосомах полностью прекращается. Из-за инактивации хромосомных генов исчезают ядрышки и постепенно разрушается ядерная оболочка. С внутренней поверхностью внутренней мембраны ядра связана тонкая пластинка (ядерная ламина) белковой природы. Эта пластинка образована промежуточными филаментами. Ядерная ламина разрушается путем деполимеризации промежуточных филаментов, а сама ядерная мембрана распадается на мелкие пузырьки. Аналогичный процесс происходит в цитоплазме: происходит уменьшение количества гранулярного эндоплазматического ретикулума, он распадается на короткие цистерны и вакуоли, количество рибосом на его мембранах резко падает. Значительно редуцируется число полисом как на мембранах, так и в гиалоплазме, что определяет общее падение синтеза белка в делящихся клетках. Аппарат Гольджи также распадается на мелкие пузырьки. В профазе также происходит образование веретена деления. Веретено деления образуется с участием центриолей и без них. Без центриолей веретено деления образуется у клеток высших растений и некоторых простейших. У некоторых других простейших и низших грибов образование веретена может проходить внутри ядра; в этом случае ядерная оболочка во время митоза не разрушается (закрытый митоз). В животных клетках главная роль в образовании веретена деления отводится центриолям. Центриоли расходятся к противоположным концам клетки и формируют полюса веретена деления. От центриолей к центромерным участкам хромосом отходят микротрубочки веретена деления. Метафаза часто занимает около трети времени всего митоза. Во время метафазы путем полимеризации белка тубулина завершается формирование веретена деления, а максимально спирализованные хромосомы выстраиваются в экваториальной плоскости клетки, и образуют так называемую метафазную пластинку (материнскую звезду). В состав веретена деления входят микротрубочки трех типов: кинетохорные (связывают каждую хроматиду с одной из центриолей. Кинетохор – специальный белковый комплекс в области центромеры), полярные (идут от одной из диплосом к центру веретена, где перекрываются с микротрубочками от другого полюса), астральные (направлены от центромеры к поверхности клетки). Постепенно в хромосомах разрушаются когезиновые комплексы между сестринскими хроматидами. К концу метафазы завершается процесс обособления друг от друга сестринских хроматид. Их плечи лежат параллельно друг другу, и соединяются только в зоне центромер. Анафаза начинается внезапно. Это самая короткая стадия митоза. Хромосомы теряют центромерные связки и синхронно начинают удаляться друг от друга по направлению к противоположным полюсам клетки. При этом они ориентированы центромерными участками к соответствующему полюсу, а теломерными – к экватору клетки. Движение хромосом осуществляется за счет сокращения микротрубочек. Происходит укорочение (разборка) кинетохорных микротрубочек и удлинение полярных микротрубочек (что ведет к расхождению самих полюсов). Кроме того, в процессе участвуют белкитранслокаторы. Они перемещают хромосомы вдоль кинетохорных микротрубочек и перемещают полярные микротрубочки в стороны друг от друга. Телофаза начинается с остановки хромосом и кончается началом реконструкции нового интерфазного ядра и разделением исходной клетки на две дочерние (цитокинез). С хромосомами ассоциируют пузырьки, образовавшиеся в профазе из разрушенных ядерных мембран. В их стенки вновь встраиваются комплексы ядерных пор. Через поры в пузырьки проникают белки, формирующие промежуточные филаменты, которые в свою очередь, образуют ядерную ламину. Благодаря этому пузырьки сливаются. Вначале они образуют двойную оболочку вокруг каждой хромосомы. Получаются мини-ядра (кариомеры). Позднее сливаются сами кариомеры, связанные с одной диплосомой. Хромосомы постепенно деконденсируются, и начинают формироваться ядрышки. В телофазе происходит разрушение митотического веретена деления. Цитокинез (цитотомия) у растительной клетки происходит путем внутриклеточного образования клеточной перегородки, а у клеток животных – путем перетяжки, впячивания плазматической мембраны внутрь клетки. Для разделения клетки по экватору формируется актомиозиновое кольцо, которое постепенно сжимается, стягивая за собой плазмолемму и образуя перетяжку все уменьшающегося диаметра. Регуляция клеточного цикла Ключевую роль в смене фаз клеточного цикла играют специальные ферменты – протеинкиназы. Это т.н. циклинзависимые киназы (Cdk – cyclindependent kinases). Каждая из них фосфорилирует определенные белки, вовлеченные в соответствующую фазу цикла, они меняют свою конфигурацию и таким образом, активируют или инактивируют их. Молекула Cdk сама по себе не активна, для ее активации требуется связывание с ней специального белка – циклина (Ц). Название «циклины» обусловлено цикличным изменением концентрации данного белка в течение клеточного цикла. При взаимодействии образуется активный комплекс циклин-Cdk. Существует несколько видов циклинов и несколько видов Cdk (разные циклины обозначаются латинскими буквами, а разные Cdk – арабскими цифрами). Комплекс циклин- Cdk очередной стадии цикла должен обеспечить: 1. Инактивирование комплекса предыдущей стадии; 2. Стимуляцию событий своей стадии; 3. Образование или активацию комплекса следующей стадии. Запускают цикл комплексы ЦД(1-3)-Cdk4 и 6. Они функционируют на начальной стадии постмитотического (G1) периода и вызывают соответствующие внутриклеточные события (рост клетки, синтез белков для репликации). Для осуществления митотического деления необходим внешний стимул – действие специфических факторов роста (митогены). 1. 2. 3. 4. 5. Известно несколько факторов роста: PDGF – фактор роста из тромбоцитов. Стимулирует пролиферацию клеток соединительной ткани. Освобождается при разрушении сосудистой стенки, участвует в процессах тромбообразования и заживления ран. EGF (эпидермальный фактор роста) – стимулирует размножение многих типов клеток; работает как сигнальный белок при эмбриональном развитии. NGF – фактор роста нервов. Осуществляет рост отдельных типов нейронов. Эритропоэтин – осуществляет пролиферацию предшественников эритроцитов. Интерлейкины (IL) – Участвуют в реакциях клеточного иммунитета и кроветворения, где выступают как стимуляторы и ингибиторы размножения и дифференцировки лимфоцитов на разных этапах иммунного ответа (лимфокины). Есть также и антимитогенные факторы (например, фактор некроза опухолей), которые останавливают деление опухолевых клеток. Факторы роста узнают соответствующий рецептор на поверхности клетки, в результате чего активируется ряд специфических ферментов – митогенактивируемых протеинкиназ, что приводит к фосфорилированию ряда транскрипционных факторов, в результате чего они активируются и запускают транскрипцию ряда генов. Среди таких генов находятся гены циклина D, Cdk4,6 – компоненты комплексов первой половины G1периода. На этих генах синтезируются соответствующие белки, которые образуют активные комплексы G1-периода. Количество Cdk в клетках регулируется присоединением специфических ингибиторов. Действие комплексов G1 периода. Комплексы циклин D-Cdk4,6 действуют на несколько субстратов: Во-первых, белок pRb (от «ретинобластома» - при мутации этого гена развивается соответствующая опухоль). Функция белка pRb состоит в ингибировании комплекса транскрипционных факторов (E2F и DP). В неделящихся клетках белок pRb находится в нефосфорилированном состоянии, поэтому имеет высокое сродство к комплексам транскрипционных факторов и блокируют их. Под действием ЦD-Cdk4,6 pRb фосфорилируется, в результате чего теряет сродство к сответствующим транскрипционным факторам (т.е. они освобождаются), в результате чего они активируется и запускают работу ряда генов: – Гены ключевых ферментов репликации ДНК (например, ДНКполимеразы а). Это неодходимо для подготовки клетки к репликации; – Предположительно гены транскрипционных факторов, стимулирующих в конце G1-периода экспрессию гена циклина В. – Гены компонентов комплексов последующих стадий цикла (циклинов Е и А, Cdk2, Cdk1, и предположительно гены циклина В). Данные белки сразу связываются с ингибиторами (KIP) что предохраняет клетку от преждевременного начала репликации. Во-вторых, ЦD-Cdk4 действует на белок АРС (anaphase-promoting complex) - анафазу обеспечивающий фактор. Фактор АРС – это фермент приводит к быстрому распаду циклина В. Разрушение циклина В необходимо в конце метафазы митоза, для осуществления анафазы. ЦD-Cdk4 фосфорилирует АРС, инактивирует его, что обеспечивает возрастание концентрации циклина В до конца метафазы митоза. В конце G1-периода ингибиторы фосфорилируются (вероятно комплексом ЦЕ-Cdk2), что лишает их сродства к комплексам S-фазы (ЦА-Cdk2 и ЦВ-Cdk2) и клетка входит в S-период. В конце G1-периода клетка проходит т.н. «точку рестрикции». Если подготовка к митозу зашла слишком далеко, клетка входит в S-период даже в том случае, если прекратилось действие митогенов. Т.о. в данный момент клеткой принимается решение о продолжении делений или о выходе из цикла (G0). Выход из цикла может быть обратимым (стволовые клетки, фибробласты, лимфоциты), и необратимым – в случае сильной дифференцировки клеток. Действие комплексов S- и G2-периодов. Комплексами S-периода являются ЦА-Cdk2 и ЦВ-Cdk2, комплекс G2периода – ЦВ-Cdk1. Основная задача этих комплексов – обеспечить прохождение репликации т.о., чтобы каждый участок ДНК был реплицирован только один раз. На хромосомах эукариот имеется много точек начала репликации (ТНР). С каждой ТНР комплекс ферментов репликации должен связаться только один раз. В состав репликативного комплекса (РК) входит 15-20 белков. Часть этих белков образуется под влиянием комплексов G1- периода (РК2), другая часть синтезируется независимо от Cdk (РК1), но могут регулироваться путем фосфорилирования и дефосфорилирования. В самом конце G1-периода путем освобождения от ингибиторов активируются комплексы S-фазы, в т.ч. ЦА-Cdk2. Этот комплекс фосфорилирует белки репликации. Изменение конфигурации этих белков активирует их и репликативный комплекс начинает работать. При этом в фосфорилированном состоянии репликативный комплекс не может повторно связаться с ТНР. Кроме репликации во время S- и G2-периодов должно произойти еще: – Образование MPF (митоз стимулирующий фактор – mitosis-promoting factor). MPF представляет собой комплекс циклин В-Cdk1 (ЦВ и Сdk1 образуются в G1 периоде) – Торможение активности этого комплекса, во избежание преждевременного начала митоза. Это осуществляется, вероятно, путем ингибиторного фосфорилирования специальной протеинкиназой. – В S-фазе в цитоплазме происходит удвоение центриолей. В G2-периоде – синтез тубулина для образования веретена деления. Профаза и метафаза митоза: действие MPF. В про- и метафазе митоза ключевую роль играет высокая концентрация MPF. Напротив – в анафазе и телофазе решающее значение имеет низкое содержание в клетке MPF. В профазе и метафазе решающее значение имеют следующие процессы: 1. Конденсация хромосом. MPF фосфорилирует гистон Н1 и другие белки, поддерживающие структуру конденсированных хромосом. Данные белки изменяют свою конфигурацию таким образом, что становятся способными связываться с ДНК с образованием метафазных хромосом (конденсация). Данный процесс идет с затратой энергии. 2. Распад ядерной оболочки. Целостность ядерной оболочки поддерживается ядерной ламиной (тонкая сетевидная пластинка, образованная промежуточными филаментами, и прилегающая к внутренней стороне внутренней ядерной мембраны. MPF катализирует фосфорилирование остатков серина промежуточных филаментов, в результате чего конструкция «рассыпается». В результате мембрана распадается на микропузырьки. 3. Распад других мембранных структур. Мембранные структуры клетки мешают расхождению хромосом и разделению цитоплазмы, поэтому подвергаются разрушению. Вероятно это происходит путем фосфорилирования MPF структурообразующих белков, связанных с мембраной. Результатом изменения конфигурации последних является их разрушение. 4. Формирование веретена деления. Фосфорилирование MPF тубулина приводит к полимеризации его с образованием микротрубочек. 5. Предупреждение преждевременной цитотомии. В районе экватора клетки располагаются нити актина и миозина. В результате их взаимодействия и сокращения происходит разделение клетки. MPF в ранней профазе фосфорилирует легкие цепи миозина, в результате он не может реагировать с актином и разделения цитоплазмы не происходит. Анафаза и телофаза митоза. Для прохождения анафазы необходимо разрушение MPF. Это осуществляется фактором, обеспечивающим анафазу (АРС – синтезируется в G1 периоде). В метафазу митоза MPF фосфорилирует помимо других белков и APC, который активируется и начинает разрушать молекулы циклина В, входящего в состав MPF, и MPF разрушается. Для расхождения хроматид необходимо разрушение связывающих их белков (сложные мультибелковые комплексы). АРС «метит» эти белки, и они быстро разрушаются, в результате чего происходит расхождение хроматид. В делящихся клетках всегда синтезируется протеинфосфатаза. После снижения концентрации MPF концентрация протеинфосфатазы увеличивается, в результате фосфорилированные в про- и метафазе белки дефосфорилируются. Это приводит: – К восстановлению ядерной оболочки. Дефосфорилированные белки ламины склонны к полимеризации с образованием промежуточных филаментов. Центром объединения служат пузырьки, связанные с кинетохорами хромосом. Сначала образуются микроядра (кариомеры) – пузырьки с одной хромосомой внутри.Далее такие кариомеры сливаются с образованием ядра. – Деконднсации хромосом. Дефосфорилирование гистонов и др. белков способствует деконденсации хромосом. – Цитокинезу. Дефосфорилирование легких цепей миозина приводит к их взаимодействию с актином и разделению дочерних клеток. Контрольные точки. В ходе цикла клетка осуществляет самоконтроль собственного состояния. Для этого в цикле существует несколько контрольных точек. Главное, что подвергается контролю – это состояние генетического материала. В зависимости от результатов «проверки» выбирается один из вариантов дальнейших действий: – Безостановочный переход к следующей фазе цикла; – Более или менее длительная задержка на текущей стадии – для исправления обнаруженных дефектов, если такое возможно. – Запуск механизма апоптоза (программируемой клеточной смерти), если нарушения неисправимы. 1. Контрольная точка G1-периода. Остановка цикла осуществляется в случае обнаружения двухцепочечных разрывов в ДНК, неправильной сегрегации хромосом или разрушение системы микротрубочек. 2. контрольная точка S-периода. Остановка цикла осуществляется в случае недостатка нуклеотидов в клетке. Контрольная точка G2-периода. Остановка цикла в случае незавершенности репликации каких-либо участков хромосом, либо крупных повреждений ДНК, оставшихся с прошлого периода. 4. Контрольная точка метафазы митоза. Остановка цикла осуществляется в случае неправильной сборки веретена деления. 3. Предполагается, что двухцепочечные разрывы узнаются специальной протеинкиназой. В большинстве случаев хромосомных повреждений центральную роль в остановке цикла играет белок р53. Он синтезируется в клетке постоянно, но очень быстро разрушатся. При наличии в клетке хромосомных повреждений ДНК-протеинкиназа фосфорилирует р53, он становится активным. Активный р53 является транскрипционным фактором для гена белка р21, который является ингибитором всех комплексов циклин-Cdk. По этой причине происходит остановка цикла, в каком бы его периоде не находилась клетка. Если повреждения хромосом достаточно велики и их исправление затягивается, то длительно сохраняющий активность р53 начинает стимулировать как транскрипционный фактор группу генов, запускающих апоптоз. В результате в клетке начинает выполняться сложная программа самоуничтожения. Дефектная клетка распадается на фрагменты и фагоцитируется соседними клетками. Несмотря на большое количество защитных механизмов (ферменты репарации, контроль за процессами митоза, апоптоз и др.), они не являются стопроцентными. При определенных условиях возможно образование клеток с дефектным генотипом. В клетке может оказаться несбалансированный набор хромосом (в случае нерасхождения хромосом за счет нарушения веретина деления); могут образовываться полиплоидные клетки (в результате неразделения цитоплазмы); кроме того, в клетке могут присутствовать различные мутации. В случае модифицирования генов, ответственных за процессы клеточного деления, клетка может потерять контроль над делением и превратиться в опухолевую. К онкогенезу могут иметь отношение 120-150 генов человека и некоторое количество вирусных генов. Мейоз. При оплодотворении происходит процесс слияния гамет, что обеспечивает увеличение вдвое количества ДНК и хромосом в зиготе и всех клетках развивающегося организма. Поэтому, при образовании половых клеток должен существовать механизм уменьшения числа хромосом, компенсирующий их удвоение при оплодотворении. Это осуществляется в процессе мейоза. Мейоз – (от греч. Meiosis-уменьшение) деление созревания,особый способ деления клеток в результате которого происходит редукция (уменьшение) числа хромосом и переход клеток из диплоидного состояния в гаплоидное, основное звено гаметогенеза. Этот процесс состоит из двух следующих друг за другом делений, сопровождающихся лишь одним удвоением ДНК. Для мейоза характерна рекомбинация генетического материала, обмен участками между гомологичными хромосомами (кроссинговер), и активация транскрипции в профазе первого деления мейоза. I деление мейоза. Профаза I. Состоит из пяти стадий: лептотена – стадия тонких нитей, зиготена – стадия сливающихся нитей, пахитена – стадия толстых нитей, диплотена – стадия двойных нитей. Лептотена. Морфологически напоминает раннюю профазу митоза. Хромосомы очень тонкие, двойные, но сестринские хроматиды различить очень трудно. Характерно появление на хромосомах сгустков хроматина – хромомеров, они нанизаны в виде бусинок и располагаются по всей длине хромосомы. Число и расположение хромомеров характерно для каждой хромосомы. Зиготена - стадия прохождения конъюгации гомологичных хромосом. Гомологичными называют хромосомы. Имеющие одинаковую форму и размер, но одна из них получена от матери, а другая от отца (содержат одинаковый набор генов). Гомологичные хромосомы сближаются и образуют бивалент. Биваленты – парные соединения удвоенных гомологичных хромосом, т.е. каждый бивалент состоит из четырех хроматид. Число бивалентов равно гаплоидному набору хромосом. В отличие от митоза для профазы мейоза характерен синтез ДНК. На стадии зиготены синтезируется небольшое количество специфической ДНК (z ДНК). Такая ДНК состоит из уникальных последовательностей, и распределена небольшими участками по длине всех хромосом. Предполагается, что специфические по строению участки zДНК на гомологичных хромосомах узнают друг друга и на некоторое время образуют связи. Позднее эта связь осуществляется с помощью синаптонемального комплекса (тяжи белковой природы). Пахитена – стадия толстых нитей. В результате конъюгации хромосомы увеличиваются в толщине. Число толстых пахитенных хромосом гаплоидно (1n). Так как пахитенные хромосомы состоят из двух гомологов, каждый из которых состоит из двух хроматид, количество ДНК равно 4с, а число хроматид 4n. На стадии пахитены происходит чрезвычайно важное событие – кроссинговер. Кроссинговер – взаимный обмен идентичными участками между гомологичными хромосомами (I эффект мейоза). Генетическим следствием кроссинговера является рекомбинация сцепленных генов, образуются хромосомы, отличные от исходных по набору генов. В пахитене также происходит синтез небольшого количества ДНК. Этот синтез репаративен. Кроме того, в пахитенной стадии начинается активация транскрипционной активности хромосом. В это время в женских половых клетках происходит активация рибосомных генов. Диплотена – стадия двойных нитей. На этой стадии происходит взаимное отталкивание гомологов друг от друга. Сестринские хроматиды становятся различимыми. По мере отталкивания хромосом становятся видны хиазмы – места перекреста и сцепления хромосом. На стадии диплотены хромосомы приобретают вид «ламповых щеток». Хромосомы в определенных участках приобретают вид парных петель. Петли представляют собой деконденсированные участки активного хроматина. Они содержат большое количество м РНК, которая здесь синтезируется. Наличие активных хромосом в диплотене резко отличает митоз от мейоза, где, начиная с профазы, полностью прекращается синтез РНК. Активация транскрипции в пахитене и диплотене совпадает с ростом формирующихся половых клеток, особенно ооцитов. В это время клетка интенсивно синтезирует и запасает белки, необходимые для обеспечения ранних стадий развития зародыша. Диакинез – стадия обособления двойных нитей. Характеризуется уменьшением числа хиазм, укорочением бивалентов, потерей ядрышек. Это переходная стадия к собственно делению клетки. На стадии метафазы I деления мейоза биваленты выстраиваются по экватору клетки. В анафазе I деления происходит расхождение хромосом. В отличие от митоза, расходятся не сестринские хроматиды, а гомологичные хромосомы, состоящие из двух сестринских хроматид. Распределение гомологов по клеткам совершенно случайное, при этом происходит перекомбинация хромосом из разных пар (II эффект мейоза). Образовавшиеся хромосомные наборы содержат по 2с ДНК и по 2n числа хроматид. Телофаза I завершает первое деление образованием двух дочерних клеток. За первым делением следует короткая интерфаза, во время которой не происходит синтеза ДНК, и клетки приступают ко второму делению, которое по морфологии и последовательности не отличается от митотического деления. Второе деление мейоза состоит из профазы II, метафазы II, анафазы II, и телофазы II. Во время анафазы второго деления в дочерние клетки расходятся сестринские хроматиды (III эффект мейоза – независимое расхождение хроматид). Таким образом, при втором мейотическом делениии клетка 2n2с дает начало двум клеткам с гаплоидным содержанием ДНК и хромосом (nc). В результате всего процесса мейоза после двух делений из одной клетки образуются четыре гаплоидных, каждая из которых отличается по своей генетической конституции. Таким образом, биологический смысл мейоза заключается в поддержании постоянства числа хромосом у разных поколений организмов, размножающихся половым путем. Половое размножение включает стадию оплодотворения – слияния двух гамет. Без механизма редукции в мейозе число хромосом удваивалось бы в каждом поколении. Кроме того, во время мейоза образуется большое количество гамет, различающихся по набору хромосом. Генетическое разнообразие обеспечивается кроссинговером и независимым комбинированием хромосом и хроматид. VII. Гаметогенез. Гаметогенез – процесс образования половых клеток. Процесс образования женских половых клеток называется оогенезом, мужских – сперматогенезом. Гаметогенез подразделяется на ряд стадий: размножения, роста, созревания и формирования. Диплоидные клетки, из которых образуются гаметы, называются сперматогониями и овогониями. Стадия размножения представляет собой ряд последовательных митотических делений сперматогоний и овогоний, в результате чего возрастает их количество. Сперматогонии размножаются на протяжении всего периода половой зрелости мужского организма. Размножение овогоний происходит в основном в период эмбриогенеза. В женском организме этот процесс наиболее интенсивно протекает между 3-м и 7-м месяцами эмбрионального развития и завершается на 3-м году жизни. На стадии роста происходит увеличение клеточных размеров и превращение сперматогониев и овогониев в сперматоциты и овоциты I порядка. Основным событием стадии созревания является мейоз. После первого деления мейоза образуются спематоциты и овоциты II порядка, после второго – сперматиды и зрелая яйцеклетка. Каждый сперматоцит II порядка дает 4 сперматиды, а каждый овоцит II порядка – 1 полноценную яйцеклетку и 3 редукционных направительных тельца, не участвующих в размножении. Благодаря этому в яйцеклетке концентрируется максимальное количество питательных веществ. Процесс сперматогенеза завершается стадией формирования (спермиогенеза). Ядра спематид уплотняются вследствие гиперспирализации хромосом, которые становятся функционально инертными. Комплекс Гольджи перемещается к одному из полюсов ядра, образуя акросому, играющую важную роль в оплодотворении. Центриоли располагаются у противоположного полюса ядра, от одной из них отрастает жгутик, у основания которого концентрируются митохондрии. На этой стадии почти вся цитоплазма сперматиды отторгается, зрелый сперматозоид практически лишен ее.