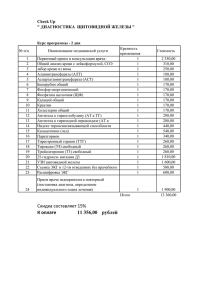
Автореферат - Дальневосточный государственный аграрный
реклама

На правах рукописи Силкин Иван Иванович ВОЗРАСТНЫЕ И СЕЗОННЫЕ СТРУКТУРНО-ФУНКЦИОНАЛЬНЫЕ ПЕРЕСТРОЙКИ НЕКОТОРЫХ ПОЛОВЫХ, ЭНДОКРИННЫХ И МУСКУСНЫХ ПРЕПУЦИАЛЬНЫХ ЖЕЛЕЗ САМЦОВ ОНДАТРЫ 06.02.01 Диагностика болезней и терапия животных, патология, онкология и морфология животных АВТОРЕФЕРАТ диссертации на соискание ученой степени доктора биологических наук Благовещенск - 2013 Работа выполнена в Федеральном государственном бюджетном образовательном учреждении высшего профессионального образования «Иркутская государственная сельскохозяйственная академия» и Федеральном государственном бюджетном образовательном учреждении высшего профессионального образования «Бурятская государственная сельскохозяйственная академия им. В.Р. Филиппова» Министерства сельского хозяйства Российской Федерации. Научный консультант доктор ветеринарных наук, профессор Попов Александр Петрович Официальные оппоненты: Сиразиев Ромазан Закарьянович доктор биологических наук, профессор, Федеральное государственное учреждение «Научно-исследовательский институт ветеринарии Восточной Сибири» Сибирского отделения Российской академии сельскохозяйственных наук, заместитель директора по научной работе Овчаренко Нина Дмитриевна доктор биологических наук, профессор, Федеральное государственное бюджетное образовательное учреждение высшего профессионального образования «Алтайский государственный университет, заведующая кафедрой зоологии и физиологии человека и животных Донкова Наталья Владимировна доктор ветеринарных наук, профессор, Федеральное государственное бюджетное образовательное учреждение высшего профессионального образования «Красноярский государственный аграрный университет», заведующая кафедрой анатомии и гистологии животных Ведущая организация Федеральное государственное бюджетное образовательное учреждение высшего профессионального образования «Якутская государственная сельскохозяйственная академия Защита состоится «30» мая 2013 года в 10:00 часов на заседании диссертационного совета Д 220.027.02 при ФГБОУ ВПО «Дальневосточный государственный аграрный университет» на факультете ветеринарной медицины и зоотехнии в ауд. 14(а) по адресу: 675005 Амурская область, г. Благовещенск, ул. Политехническая 86, факс: (416-2) 52-62-80, Е-mail: dalgau@tsl.ru. С диссертацией можно ознакомиться в библиотеке ФГБОУ ВПО «Дальневосточный государственный аграрный университет», на сайте ВАК РФ http//vak.ed.gov.ru и на сайте ДальГАУ http: //www.dalgau.ru Автореферат разослан «12» марта 2013 года Ученый секретарь диссертационного совета Самусенко Ольга Леонидовна 3 ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ Актуальность темы. В отечественной и зарубежной литературе в последнее время отмечается заметное повышение интереса к всестороннему изучению органов эндокринной регуляции животных и человека. Это объясняется рядом положений. Прежде всего, следует отметить, что их гормоны – влияют на многие биологические процессы (обмен веществ, терморегуляцию, рост и размножение, функциональную активность всех органов и систем, ЦНС и др.) Известна высокая лабильность функциональной активности этих органов при различных воздействиях. Показано, что они принимают участие в реакциях адаптации. Значение эндокринной системы ставит эндокринологию на одно из ведущих мест среди биологических наук. Повышенное внимание со стороны ученых, медицинских работников, биологов к проблемам бесплодия, безусловно, привело к развитию андрологического направления не только в медицине, но и в ветеринарии (Попов А.П., 2004). Выбор этого направления оказался весьма своевременным и перспективным, так как на сегодняшний день в медицине сформировалась новая область науки: инфлогенитология, которая изучает взаимосвязь репродуктивной системы с другими органными системами и в первую очередь с эндокринной (Ромашова М.Ф. и др., 1998). Помимо практического значения изучения внутренних половых органов самцов и эндокринных органов, результаты наших исследований могут быть использованы в эколого-гистологических исследованиях. Этот термин был предложен Ю.Н. Олейником (2003), изучавшем в этом аспекте щитовидную железу крапчатого суслика. Среди всех видов охотничье-промысловых животных ондатре принадлежит пальма первенства по величине ареала, искусственно созданного человеком. Экономическая эффективность акклиматизации и искусственного расселения этого грызуна не вызывает сомнения. Кроме того, с ее помощью удалось решить проблему утилизации водно-болотной и прибрежно-водной растительности на водоемах нашей страны (Ширяев В., 1989). Данный вид мелких млекопитающих благодаря многочисленности, широте распространения и удобству изучения может успешно использоваться в разнообразных экологических исследованиях в качестве тест-объекта. Метод морфофункциональных индикаторов часто используется в эколого-физиологических исследованиях. Он позволяет более объективно оценить реакцию организма на конкретные природно-климатические факторы и в условиях техногенных загрязнений. Изменчивость морфофункциональных характеристик органов репродукции и эндокринной регуляции может быть связана с определенными адаптивными сдвигами метаболизма животных при действии разнообразных факторов окружающей среды. 4 Следует также отметить, что территория экосистемы озера Байкал относится к биогеохимическим провинциям, характеризующиеся дефицитом йода в биосфере и наличием среди населения эндемического зоба (Муратова Н.М., 2002). Помимо исследования органов репродукции и эндокринной регуляции у самцов ондатры интересным объектом исследования явился специфический орган – мускусные препуциальные железы. В доступной нам литературе есть единичные работы по этому органу, и то они выполнены на взрослых животных (Reisinger L., 1916, Schaffer J., 1940, Соколов В.Е. и др., 1993). В брошюрах и книгах об ондатре мускусные препуциальные железы либо совсем не упоминаются, либо неправильно указывается, что они имеются только у самцов. О морфофункциональных изменениях органов эндокринной регуляции и репродукции у человека и разных видов животных накоплен колоссальный материал, представленный в ряде оригинальных работ. Однако в доступной нам литературе мы не обнаружили данных о морфофункциональном состоянии органов репродукции, эндокринной регуляции и видоспецифичных мускусных препуциальных желез ондатры в постнатальном онтогенезе и при воздействии сезонных изменений окружающей среды. Впервые эти исследования проведены в природноклиматических условиях экосистемы Южного Прибайкалья. Цель и задачи исследования. Настоящая работа является самостоятельным разделом общей комплексной темы кафедры анатомии и ветеринарно-санитарной экспертизы Иркутской государственной сельскохозяйственной академии (номер государственной регистрации 01.2.00900999) и кафедры анатомии, гистологии и патоморфологии Бурятской государственной сельскохозяйственной академии им. В.Р. Филиппова (номер государственной регистрации 01.2.01156775). Целью настоящих исследований явилось изучение возрастных и сезонных морфофункциональных изменений тканевых и клеточных структур внутренних половых органов, щитовидных, надпочечных и мускусных препуциальных желез самцов ондатры, обитающих в условиях экосистемы Южного Прибайкалья. Исходя из цели исследования, необходимо было решить следующие задачи: – изучить анатомо-топографические особенности внутренних половых органов, щитовидной, надпочечных и мускусных препуциальных желез самцов ондатры; – изучить органо- и гистогенез внутренних половых органов, щитовидной, надпочечных и мускусных препуциальных желез самцов ондатры обитающих в условиях экосистемы Южного Прибайкалья; – проследить морфофункциональные изменения внутренних половых органов, щитовидной, надпочечных и мускусных препуциальных 5 желез самцов ондатры в период спада половой активности до наступления стадии полового покоя в условиях экосистемы Южного Прибайкалья; – выявить физиологическую взаимосвязь внутренних половых органов, щитовидной, надпочечных и мускусных препуциальных желез самцов ондатры Научная новизна и ценность полученных результатов заключается в том, что методами классической гистологии, современной гистохимии и биометрии в дифференциальной оценке белковых и углеводных компонентов, а также исследованиями уровня тиреоидных и андрогенных гормонов методами радиоиммунологического и иммуноферментного анализов, выявлен ряд новых закономерностей структурно-функциональной дифференциации внутренних половых органов, щитовидной, надпочечных и мускусных препуциальных желез самцов ондатры в различные периоды постнатального онтогенеза и в период спада половой активности вплоть до наступления стадии полового покоя на территории Южного Прибайкалья. Полученные данные являются оригинальными, расширяют и дополняют отдельные положения теории индивидуального развития систем органов и организма в целом. Теоретическая и практическая значимость. Полученные нами данные расширяют современные представления о биологии ондатры, особенностях развития и строения семенников и придаточных половых органов самцов, органов эндокринной регуляции и видоспецифичных органов (мускусные препуциальные железы) в условиях уникальной экосистемы Южного Прибайкалья. Обнаружен ряд особенностей в действии факторов (природно-климатические условия) на морфофункциональные показатели и состояние этих органов. Результаты гистологических и гистохимических исследований по вопросам возрастных и сезонных особенностей структуры и функций органов репродукции и эндокринной регуляции ондатры в постнатальном онтогенезе представляют несомненный интерес и могут быть использованы при написании соответствующих разделов биологии, зоологии, охотоведения и звероводства, сравнительной гистологии и гистохимии. Биометрические исследования тканевых и клеточных структур органов репродукции и эндокринных органов могут быть использованы как базовые (нормативные) при изучении влияния природно-климатических факторов на эти органы, степени техногенных загрязнений в данном регионе, учитываться для регуляции промысла и разумного использования популяций ондатры, а также могут быть использованы для прогноза влияния биогеохимических условий на состояние биоты. Результаты исследований мускусной препуциальной железы помогут глубже понять физиологические процессы, происходящие в специфическом функциональном органе самцов ондатры на различных этапах постнатального онтогенеза. 6 Более того, материалы данной работы будут служить основой для конкретизации дальнейших исследований экологии, морфологии и биологии ондатры, а также способствовать разработке региональных ландшафтногеографических принципов рационального использования ресурсов вида. Внедрение исследований. Полученные данные используются в практической работе службой по охране и использованию животного мира Иркутской области, Государственным учреждением Амурской области «Дирекция по охране и использованию животного мира и ООПТ», промыслово-охотничьем хозяйством ЗАО «Новый Промой» Кабанского района Республики Бурятия; учебной и научной работе отдела эволюционной морфологии позвоночных животных Института зоологии им. И.И. Шмальгаузена НАН Украины, научно-исследовательского центра биобезопасности и экологического контроля ресурсов АПК Днепропетровского государственного аграрного университета; кафедр факультета охотоведения Иркутской государственной сельскохозяйственной академии; кафедры анатомии, гистологии и патоморфологии Бурятской государственной сельскохозяйственной академии им. В.Р. Филиппова; кафедры морфологии, патологии и физиологии животных Дальневосточного государственного аграрного университета; кафедры анатомии и гистологии животных Красноярского государственного аграрного университета; кафедры патологической анатомии и гистологии Витебской ордена «Знак Почёта» государственной академии ветеринарной медицины; кафедры анатомии животных им. акад. В.Г. Касьяненка Национального университета биоресурсов и природопользования Украины; кафедры анатомии и физиологии животных Полтавской государственной аграрной академии; кафедры анатомии и гистологии им. П.А. Ковальского Белоцерковского национального аграрного университета; кафедры анатомии сельскохозяйственных животных Львовского национального университета ветеринарной медицины и биотехнологий; кафедры анатомии, нормальной и патологической физиологии животных Сумского национального аграрного университета. Апробация работы. Основные положения диссертации доложены на научно-практических конференциях Бурятской государственной сельскохозяйственной академии (Улан-Удэ, 2001-2009), Иркутской государственной сельскохозяйственной академии (Иркутск, 2004-2010), 5 Всероссийской конференции «Актуальные вопросы ветеринарной медицины мелких домашних животных» (Екатеринбург, 2003), 2 научно-практической конференции «Роль молодых ученых в развитии науки» (Великие Луки, 2007), Международной научно-практической конференции «Состояние и перспективы обеспечения ветеринарного благополучия Восточной Сибири» ГНУ НИИ ветеринарии Восточной Сибири СО РАСХН (Чита, 2008), Международной научной конференции посвященной 90-летию кафедры анатомии животных СПбГАВМ «Актуальные проблемы ветеринарной морфологии» (Санкт-Петербург, 2009), Международной научной 7 конференции посвященной 100-летию со дня рождения профессора Г.О. Гиммельрейха и 90-летию кафедры анатомии им. академика В.Г. Касьяненко НУБП Украины «Морфология XXI столетия» (Киев, 2010). Основные положения, выносимые на защиту. Возрастная морфофункциональная дифференциация внутренних половых органов, щитовидной, надпочечных и мускусных препуциальных желез самцов ондатры, обитающих в условиях экосистемы Южного Прибайкалья. Видовые особенности морфологических и гистохимических показателей внутренних половых органов, щитовидной, надпочечных и мускусных препуциальных желез у самцов ондатры. Видовая специфичность секретов внутренних половых органов, щитовидной, надпочечных и мускусных препуциальных желез самцов ондатры. Зависимость изменения морфофункционального состояния внутренних половых органов, щитовидной, надпочечных и мускусных препуциальных желез самцов ондатры от возраста и половой активности. Физиологическая взаимосвязь внутренних половых органов, щитовидной, надпочечных и мускусных препуциальных желез самцов ондатры. Публикации. Основное содержание работы отражено в 47 научных работах, из них 16 статей – в изданиях, рекомендованных ВАК РФ, 1 монография, получен 1 патент РФ на изобретение. Структура и объем диссертации. Диссертация изложена на 300 страницах компьютерного текста, состоит из следующих разделов: введения, обзора литературы, собственных исследований, анализа полученных результатов, выводов, практических предложений. Список использованной литературы включает 377 источников, в том числе 112 иностранных. Работа иллюстрирована 38 таблицами и 77 рисунками. ОСНОВНОЕ СОДЕРЖАНИЕ РАБОТЫ Материал и методы исследования Материалом для исследования являлись семенники, придатки семенников, предстательные, пузырьковидные, щитовидные, надпочечные и мускусные препуциальные железы, а также сыворотка крови от новорожденных, 1, 3, 5, 8, 10, 12-месячных самцов ондатры (Ondatra zibeticus) постнатального периода развития и половозрелых животных в разные периоды года (сентябрь, октябрь, ноябрь, декабрь). Мы не обнаружили у самцов ондатры бульбоуретральные (луковичные) железы. Изучаемый возрастной ряд представлен семью, функционально обоснованными возрастными периодами. Исследование сезонных изменений в изучаемых органах базировалось на знании о влиянии природноклиматических факторов в данном регионе на физиологию самцов ондатры. Материал собирали в период полевых экспедиций от условно здоровых 8 особей в южной зоне Прибайкалья, на территории Иркутского района Иркутской области по берегам Иркутского водохранилища (пойма реки Ангары) и в районе дельты реки Селенги Кабанского района Республики Бурятия. По величине вегетационного периода всю территорию Иркутской области и Республики Бурятия можно разделить на три зоны: южную, среднюю и северную. В южную зону входят южные районы Бурятии до широты города Улан-Удэ и дельты реки Селенги, а также южные районы Иркутской области до широты г. Иркутска (Комаров А.В., 1971). Здесь имеются такие ценные, наиболее продуктивные угодья с обильными запасами кормовых растений как пойма реки Ангары с Иркутским водохранилищем и дельта реки Селенги с прилегающими байкальскими сорами. Возраст определяли при помощи соответствующих методик (Корсакова И.Б., 1970; Водопьянов Б.Г., Саловаров В.О., 2001). Всего исследован материал от 198 животных. Полученный материал фиксировали в 10% растворе нейтрального формалина, жидкости Карнуа, в нейтральной смеси Шабадаш и заключали в парафин. Полученную сыворотку крови замораживали при температуре – 18°С. Осуществлен комплекс мероприятий, включающий в себя различные методы исследования: • Морфологические исследования. Общие макро-морфометрические исследования. Для оценки морфофункциональных параметров использовали животных отловленных капканами (капкан № 1). Тушки животных подвергали взвешиванию с точностью до 1,0 г. После изъятия органов из тушек, их подвергали взвешиванию с целью определения относительной массы исследуемых органов к массе тела животных в зависимости от возраста и сезона года. При осмотре отмечали цвет, консистенцию, форму и топографию. Общие микро-морфометрические исследования. Для оценки клеточных элементов и тканевых структур изготавливали серийные парафиновые срезы толщиной 5-7 мкм на санном микротоме «C. Reichert wien», с последующей их окраской гематоксилин-эозином по Караччи, железным гематоксилином по Гейденгайну и по методу Ван-Гизон. Гистологические препараты изучали при помощи микроскопов «ЛОМО Микмед-1» и «Motic BA 400 T», микро-морфометрические измерения производили при помощи винтового окуляр-микрометра МОВ-16-1. При выборе морфологических критериев оценки функциональной активности изучаемых органов использовали как общие микроморфометрические методы исследования, которые универсальны для всех исследуемых органов, так и специфические, учитывающие особенности их гистологического строения. Во всех исследуемых органах определяли индекс склерозирования – соотношение относительных объемов паренхимы и стромы. При вычислении 9 объема ядер клеток пользовались формулами для определения объемов круглых ядер (с коэффициентом элонгации менее 1,2) по формуле: V π овальных ядер по формуле: V D3 , 6 6 ( L B) 3 , где L – малый диаметр ядра, В – большой диаметр ядра (Хесин Я.Р., 1967; Ташке К., 1980). Ядерно-цитоплазматическое соотношение (ЯЦС) клеток вычисляли по формуле: d 2 ср. я ЯЦС= 2 , d ср.к где d2 ср.я – средний диаметр ядра, d2 ср.к – средний диаметр клетки (Бондаренко О.В. и др., 2000). Причем чем выше значения ЯЦС, тем активнее клетка (Диденко А.А. и др., 2006). Для оценки функциональной активности клеточных ядер, проводили окраску альциановым синим и сафранином Т (Яцковский А.Н., 1987). При этом ядерные структуры, окрашивающиеся в красный цвет, учитывались как гетерохроматин, а структуры синего цвета считали эухроматином. Специальные микро-морфометрические исследования. Семенник. В гистоструктуре семенника определяли диаметр извитых семенных канальцев, клеточный состав органа, а также стадии цикла сперматогенного эпителия (Ortavant R., 1959). Придаток семенника. Изучение придатка семенника осуществлялось в различных его частях: головке, теле и хвосте придатка. Проводили измерения диаметра протока и просвета выносящих семенных канальцев. Предстательная и пузырьковидная железа. В гистоструктуре добавочных половых желез нами изучалась высота эпителия в концевых отделах и выводных протоках желез, а также диаметр выводных протоков. При выборе морфологических критериев оценки функциональной активности половых желез руководствовались работами А.П. Попова (1981, 1995, 2004). Щитовидная железа. Для оценки морфологических критериев функциональной активности щитовидной железы исследовали следующие показатели: Форма и средний диаметр фолликулов, их число в поле зрения, процентное соотношение крупных, средних и мелких фолликулов. Объем фолликулов (исходя из представлений о фолликулах как об эллипсоидах) по формуле: V L B2 6 где L и B – больший и меньший диаметры фолликула. Среднюю площадь тиреоцитов (исходя из формы трапеции) по формуле (Дмитриева Н.И., 1990; Дубынин В.А., 1996): 10 Sk n1 n2 h 2 где S k – площадь клетки, h – высота клетки, n1 и n2 – большее и меньшее основание клетки (трапеции). Оценка функциональной активности щитовидной железы проводилась с помощью вычисления фолликулярно-коллоидного индекса (индекса Брауна), который обозначает соотношение относительных объемов фолликулярного эпителия и коллоида (функциональная активность прямо пропорциональна относительному объему эпителия и обратно пропорциональна относительному объему коллоида). При исследовании щитовидной железы руководствовались работами О.А. Гомбоевой (2005); Е.С. Горбачевой (2006); Е.С. Манюк (2008). Надпочечник. При выборе морфологических критериев оценки функционального состояния надпочечной железы исследовали следующие морфологические показатели: абсолютную толщину коркового и мозгового слоя и их соотношение, абсолютную и относительную толщину клубочковой, пучковой и сетчатой зон коры. Толщину различных зон коркового и мозгового вещества измеряли в 10 полях зрения на гистопрепарате от каждого животного. В работе по изучению надпочечника пользовались трудами П.М. Торгуна (1968); Е.С. Горбачевой (2006). Мускусная препуциальная железа. При выборе морфологических критериев оценки функциональной активности мускусной препуциальной железы руководствовались работами В.Е. Соколова и др. (1993); С.А. Шабадаш (2005); С.А. Шабадаш, О.Ф. Чернова (2006). Нами учитывались следующие морфологические и морфометрические показатели: форма и средний диаметр железистых клубочков, ширина и длина долей, высота эпителия в концевых отделах, выводных протоках, а также диаметр, просвет и количество выводных протоков. Полученные числовые данные макро- и микрометрических показателей подвергали статистической обработке с использованием компьютерных программ «Microsoft Excel», «Biostatica», при этом учитывая классические работы некоторых авторов (Плохинский Н.А., 1970; Урбах В.Ю., 1975; Лакин Г.Ф., 1980). Гистохимические исследования. Для выявления гликогена и других ШИК-положительных веществ использовали метод А.Л. Шабадаша (1947). Для дифференциации гликогена от других ШИК-положительных компонентов гистологические препараты перед окраской подвергали обработке амилазой слюны в течение 20 минут при температуре 37°С (Кононский А.И., 1976), а также в нашей модификации 0,5%-ным раствором α-амилазы (α-Amylase from Aspergillus oryze) производства фирмы «Fluka» (Швейцария) в буферном растворе при рН 5,6 в течение 20 минут при температуре 37°С. Для идентификации нейтральных гликопротеинов учитывали ШИКреакцию после предварительной обработки гистологических срезов 5%-ным 11 раствором солянокислого фенилгидразина в течение 1 часа при комнатной температуре (Spicer S.S., 1961). Кислые группы углеводных соединений выявляли основным коричневым (Шубич М.Г., 1961), альциановым синим при рН-1,0 (Lev R., Spicer S.S., 1964) и при pH-2,7 (Mowry R.W., 1956), толуидиновым синим при pH-4,6, реакцией «скрытой метахромазии» (Виноградов В.В., Потапова В.Б., 1961). Для идентификации кислых гликопротеинов использовали ферментативный контроль с бактериальной гиалуронидазой (Streptomyces hyalurolyticus) фирмы «Sigma» в концентрации 1 мг/мл на физиологическом растворе в течение 3 часов при температуре 37°С (Quintarelli G., 1963). Для выявления общего белка использовали метод тетразониевого азосочетания по Даниелли с применением прочного синего Б по Берстону (Пирс Э., 1962) в нашей модификации (Силкин И.И., Власов Б.Я., 2009), где взамен труднодоступного веронала (барбитала) мы использовали глицингидроокись натрия (Кочетов Г.А., 1980). Бромфеноловый синий применяли для обнаружения суммарного (при рН-2,2) и свободного катионного (при рН8,2) белков (Ringerts N., Zetterberg А., 1966). При выявлении последнего гистопрепараты предварительно обрабатывали 3%-ным раствором трихлоруксусной кислоты при t 90˚С с целью удаления нуклеиновых кислот и освобождения связанных с ними белков типа гистонов. РНК выявляли по методу Браше в модификации N.B. Kurnick (1955) с постановкой контроля рибонуклеазой (Ribonuclease from bovine pancreas) производства фирмы «Fluka» (Германия) в концентрации 1мг/мл на физиологическом растворе в течение 1 часа при t 37˚С. За РНК принимали вещества, проявляющие пиронинофилию, ферментирующиеся рибонуклеазой. ДНК идентифицировали стандартной реакцией Фельгена (Лилли Р.Д., 1969). Взамен трудно доступных М-проционовых красителей для выявления аминокислот лизина и гистидина мы использовали остазиновые красители (остазин – С-6Г), рекомендованные в гистохимическую практику А.П. Поповым (1981). Для обнаружения аргинина использовали реакцию с тиазиновым красным (Шубич М.Г. и др., 1975) и качественную реакцию на аргинин с 8-оксихинолином-гипохлорита (Лилли Р.Д., 1969). Идентификацию лизина, гистидина и аргинина проводили дезаминированием в свежеприготовленной смеси равных объемов 5%-ного раствора нитрита натрия и 10%-ного раствора уксусной кислоты при t +4˚С в течение 24 часов. Дезаминирование блокирует лизин, сохраняя окрашивание гистидина и аргинина. Таким образом, проведение комплекса гистохимических реакций позволило выявить большинство функциональных групп углеводных и белковых компонентов и на этом основании, более предметно судить о функциональных особенностях исследуемого эпителия и его секрета. При этом, для оценки функционального состояния органов в нашем случае у 12 диких животных, наиболее высокоинформативными методами являются морфологические и гистохимические. Гормональные исследования. Для проведения гормональных исследований от животных была получена кровь. От новорожденных кровь получали путем декапитации, от взрослых путем перерезки сонной артерии. Кровь собирали в стерильные химические стаканы, затем переливали в стерильные одноразовые пробирки производства фирмы «Vacuette» для получения сыворотки. Сыворотку крови исследовали на содержание трийодтиронина (Т3), тироксина (Т4), тестостерона, лютропина (ЛГ) и фоллитропина (ФСГ). Концентрацию гормонов в сыворотке крови определяли методами радиоиммунологического и иммуноферментного анализов с использованием коммерческих тест-наборов «Диас» (Россия) и «Алкорбио» (Россия). Полученные данные обрабатывали методами вариационной статистики и корреляционного анализа. По результатам исследований анализировали содержание указанных гормонов в зависимости от возраста и сезона года. МОРФОФУНКЦИОНАЛЬНЫЙ ГИСТОГЕНЕЗ СЕМЕННИКА Возрастные морфологические изменения семенника Нами установлено, что у самцов ондатры всех исследуемых возрастных групп имеются следующие внутренние половые органы: парные половые железы: семенники с придатками; семяпроводы с их железистой частью (ампулами семяпроводов); придаточные половые железы; к ним относятся парные: предстательная и пузырьковидная железы; у самцов ондатры отсутствуют луковичные (бульбоуретральные) железы. Семенники располагаются в тазовой полости на уровне седалищных бугров под париетальным листком брюшины и приближены к анальному отверстию, овальной формы бело-желтоватого цвета с хорошо различимым придатком. Под белочной оболочкой располагаются многочисленные кровеносные сосуды. Формирование гонады связано с увеличением ее массы. Изменения диаметра семенных канальцев находятся в прямой зависимости от изменения толщины слоя сперматогенного эпителия. Установлена тесная положительная корреляция между возрастом и диаметром семенных канальцев (r= 0,84). У новорожденных и 1-месячных щенков ондатры развитие тканевых структур гонад и их клеточного состава является далеко не завершенным. Паренхима органа представлена формирующимися семенными канальцами, где более крупные канальцы расположены по периферии, а мелкие ближе к сети семенника. Гонады покрыты белочной оболочкой с нежными оксифильными волоконцами. В семенных канальцах находятся сустентоциты и гоноциты. Ядра сустентоцитов округлой формы с четким ядрышком. 13 Между канальцами rete testis и семенными канальцами в строме гонад обнаруживаются скопления интерстициальных эндокриноцитов. Мы выделили два вида ИЭ, большую часть составляют ИЭ с округлыми и округло-овальными светлыми ядрами средних размеров со слабо оксифильной цитоплазмой, и с мелкими, светлыми ядрами, имеющими одно ядрышко. У 3-месячных самцов ондатры белочная оболочка более развита, чем у новорожденных и 1-месячных щенков, состоит из двух слоев: поверхностного, состоящего из более плотной, и внутреннего, из менее плотной соединительной ткани, она резко утолщается, с перехватами. В этом возрасте становится хорошо выраженной базальная мембрана и собственная пластинка извитых семенных канальцев. В канальцах заметны митотически делящиеся гоноциты. Увеличивается диаметр семенных канальцев и высота эпителия (табл. 1). Существенно изменяется паренхимо-стромальное соотношение, в сторону увеличения удельного объема паренхимы. Волокна и клетки вокруг терминальных отделов семенных канальцев и канальцев rete testis образуют более мощный соединительнотканный каркас, по сравнению с интерстициальной тканью, окружающей семенные канальцы. Наблюдается уменьшение количества интерстициальных эндокриноцитов в интерстиции. В 5 месяцев митотическая активность гоноцитов увеличивается, и они дают начало новой генерации –просперматогониям. В центральной части канальцев эпителий становится менее плотным, появляются участки разрежения. В 8 месяцев гонады являются хорошо сформированным органом, покрытым снаружи белочной оболочкой. Происходит начальный процесс развития половых клеток. Существенно увеличивается диаметр извитых семенных канальцев (табл. 1). Отмечается изменение высоты эпителия, которая, по сравнению с предыдущим возрастом, увеличивается почти в 2 раза (табл. 1). У животных этой возрастной группы можно встретить различные сочетания клеток, находящихся на разных стадиях мейотического деления. В некоторых канальцах обнаруживаются сустентоциты, гоноциты, просперматогонии, смерматогонии А и первичные сперматоциты. Обнаруживаются канальцы, количество которых является доминирующим, где идентифицируются сперматогонии А и В и сперматоциты, которые находятся на стадии пахитены и диплотены. В большинстве ядра половых клеток содержат эухроматин. Интерстициальная ткань несколько уменьшается. В ней обнаруживаются ИЭ средних размеров, расположенные небольшими скоплениями между извитыми семенными канальцами. В 10-12 месяцев нами отмечен активный сперматогенез и наличие 8 стадий цикла сперматогенного эпителия. Наибольшей своей величины семенные канальцы достигают в 10-12 месяцев (табл. 1). Следовательно, можно сказать, что динамика изменений массы семенников, диаметра семенных канальцев, высоты эпителия находится в прямой зависимости от возраста и годового цикла животного. Таблица 1 Морфометрические показатели семенника самцов ондатры в зависимости от возраста Показатели 1 23,5±0,25*** 22,2±0,71*** 3,0±0,24** 8,5±0,10 11,0±0,07** 49,0±2,90** 164,7±20,23** 0,5±0,01* 0,6±0,02*** 55,4±0,85** 15,2±0,20*** 20,4±0,60*** 3 26,0±1,37*** 24,8±1,32*** 3,1±0,14*** 8,6±0,06** 11,0±0,05** 51,7±2,50*** 155,4±16,60*** 0,5±0,03* 0,6±0,01 66,3±0,65*** Возраст (мес.) 5 30,3±2,70*** 32,0±1,16*** 3,0±0,18 8,8±0,11 11,0±0,20** 59,2±4,20 167,8±7,55*** 0,5±0,02** 0,6±0,01*** 76,0±1,01*** 8 45,0±0,75 44,4±1,87*** 3,0±0,20*** 8,8±0,10*** 11,0±0,30*** 59,6±1,54*** 177,2±8,12*** 0,5±0,03*** 0,6±0,03*** 124,1±1,26*** 10 48,4±5,10*** 48,0±2,36 3,3±0,24*** 9,0±0,12*** 11,4±0,10** 67,4±2,92*** 210,0±5,40*** 0,5±0,01*** 0,6±0,04** 173,6±0,71*** 12 61,4±5,10 48,7±1,35 3,7±0,24 9,5±0,06 11,4±0,07 87,3±2,93 216,8±6,53 0,6±0,03 0,6±0,01 177,7±0,97 27,4±0,70*** 40,0±0,80*** 74,8±1,04*** 84,7±0,86 87,8±1,70 Примечания: С – сустентоциты, Г – гоноциты; различия с предыдущей группой достоверны: *** – при Р<0,001; ** – при Р<0,01; * – при Р<0,05 14 Толщина капсулы (мкм) Кол-во сустентоцитов Кол-во гоноцитов Средний диаметр С клеток (мкм) Г Объем ядер клеток С (мкм3) Г ЯЦС (ед.) С Г Диаметр извитых семен. канальцев (мкм) Высота эпителия (мкм) Новорожден 23,0±0,64 22,1±0,38 3,1±0,11* 8,3±0,07*** 11,0±0,10 43,1±1,80*** 167,8±5,07* 0,5±0,01 0,6±0,02 55,2±1,47 15 Возрастные гистохимические показатели семенника Характерной особенностью для возрастных изменений является выявление гликогена с 3-месячного возраста, который постепенно накапливается до 10-месячного возраста. Нейтральные гликопротеины в эпителии извитых семенных канальцев начинают выявляться с 10-месячного возраста, до этого периода они обнаруживаются в волокнах белочной оболочки, собственной пластинке семенных канальцев, в волокнах соединительной ткани. Происходит сезонное уменьшение вышеперечисленного компонента. Количество общего и катионного белков, РНК, лизина, гистидина в цитоплазме, а ДНК и аргинина в ядрах сустентоцитов увеличивается с возрастом животных и отмечается наибольшее количество в 10 месячном возрасте (половое созревание), что свидетельствует о начале белоксинтезирующей функции гонад. Гистохимическим изменениям подвергаются и интерстициальные эндокриноциты. Наличие гликогена в цитоплазме ИЭ у новорожденных самцов ондатры. С возрастом количество гликогена снижается, и только некоторые интерстициальные эндокриноциты содержат этот компонент. Выраженная интенсивная реакция на общий и катионный белки, лизин, гистидин и РНК в цитоплазме ИЭ показывает на наличие белкового синтеза. С возрастом в собственной пластинке, соединительной ткани, которая окружает канальцы rete testis и терминальные отделы, накапливается значительное содержание нейтральных гликопротеинов, сульфатированных протеогликанов, общего и катионного белков, РНК, лизина и гистидина. Возрастная гормональная активность семенника Концентрация половых гормонов у новорожденных щенков ондатры была невысокой (табл. 2). В 1-3 месяца постнатального онтогенеза концентрация половых гормонов незначительно возрастает. Активация синтеза тестостерона, лютропина и фоллитропина отмечается у 5 месячных самцов (табл. 2). Высокая концентрация исследуемых гормональных показателей отмечена в 8-месячном возрасте (табл. 2). В дальнейшем, начиная с 10-месячного возраста, происходит увеличение концентрации половых гормонов в сыворотке крови. В этом возрастном периоде гормональный фон гонад самцов ондатры максимальный. В 12 месяцев концентрация половых гормонов в сыворотке самцов ондатры существенно не отличается от предыдущего периода постнатального онтогенеза (табл. 2). 16 Для выявления взаимосвязи между концентрацией гормонов в сыворотке крови и массой тела самцов ондатры были рассчитаны коэффициенты корреляции. Таблица 2 Концентрация половых гормонов в сыворотке крови самцов ондатры в зависимости от возраста Возраст (мес.) новорожденные 1 3 5 8 10 12 Тестостерон (нМ/л) 2,11±0,004*** 4,56±0,19*** 9,94±0,10*** 19,75±0,23*** 52,06±2,06*** 72,58±2,15 73,94±1,92 ЛГ (МЕД/л) 0,04±0,004 0,048±0,003*** 0,128±0,008*** 0,272±0,022*** 0,42±0,035*** 0,62±0,01** 0,94±0,11 ФСГ (МЕД/л) 0,026±0,003 0,032±0,003*** 0,044±0.002 0,046±0,002*** 0,064±0.003*** 0,84±0,11*** 0,90±0,30 Примечание: различия с предыдущей группой достоверны: *** – при Р<0,001; ** – при Р<0,01; * – при Р<0,05 Положительная корреляция между концентрацией тестостерона и массой тела выявлена у 1-месячных щенков (соответственно r= 0,32 и r= 0,35). Как известно, лютропин и фоллитропин являются гонадотропными гормонами гипофиза и воздействуя на семенники или яичники, стимулируют секрецию тестостерона или эстрадиола, определяя таким образом нормальную репродуктивную способность вида. В этой связи с целью определения возрастной взаимосвязи между концентрацией тестостерона и гонадотропных гормонов в крови самцов ондатры были рассчитаны коэффициенты корреляции. Была установлена сильная корреляционная связь между концентрацией тестостерона и лютропина (r= 0,95), а также между концентрацией тестостерона и фоллитропина (r= 0,86). СЕЗОННЫЕ МОРФОФУНКЦИОНАЛЬНЫЕ ИЗМЕНЕНИЯ СЕМЕННИКА Сезонные морфологические изменения семенника Снижение половой активности ондатры приходится на конец сентября. Преобладающее число семенных канальцев у половозрелых животных находится на 6-7 стадиях цикла, что свидетельствует о спаде активности сперматогенной функции гонад. В конце октября семенники у самцов ондатры дряблые, канальцы суживаются. Клеточный состав сперматогенного эпителия представлен сустентоцитами, гоноцитами, просперматогониями, сперматогониями А и 17 первичными сперматоцитами. В просветах семенных канальцев обнаруживаются своеобразные скопления из сперматозоидов, которые как бы склеены между собой. При окраске на функциональную активность ядер обнаружено, что большее количество клеток имеют неактивные ядра. В конце ноября в гонадах половозрелых животных просветы в семенных канальцах отсутствуют. Эпителий канальцев представлен сустентоцитами, количество которых увеличивается, и сперматогониями. Сперматогонии в дальнейшем не дифференцируются, они не переходят в свою дальнейшую стадию – первичные сперматоциты. Отдельные извитые семенные канальцы проявляют признаки дегенерации. Происходит уменьшение исследуемых морфометрических показателей семенника (табл. 3). К концу декабря у половозрелых самцов ондатры существенно снижаются все морфометрические показатели (табл. 3), что связано с наступлением полового покоя. Эпителий семенных канальцев представлен сустентоцитами и сперматогониями. Таблица 3 Морфометрические показатели семенника самцов ондатры в зависимости от половой активности Показатели Сентябрь 27,6±1,23*** 55,7±1,81*** 3,8±0,14*** С 9,4±0,11*** Г 11,4±0,16 С 82,4±1,30*** Г 212,0±1,00*** С 0,6±0,02*** Г 0,6±0,01*** Диаметр извитых семенных 171,7±9,01*** канальцев (мкм) Высота эпителия (мкм) 83,3±3,94* Толщина капсулы (мкм) Кол-во сустентоцитов Кол-во гоноцитов Средний диаметр клеток (мкм) Объем ядер клеток (мкм3) ЯЦС (ед.) Период времени (мес.) Октябрь Ноябрь 28,3±1,31*** 30,4±1,14*** 43,8±1,57*** 24,3±0,85*** 3,3±0,24 3,3±0,24** 8,8±0,10*** 8,4±0,12*** 11,0±0,10*** 10,4±0,14*** 58,4±1,00*** 54,3±0,64*** 179,4±1,92*** 164,5±0,64*** 0,5±0,03** 0,5±0,01*** 0,6±0,03 0,6±0,03** 113,4±5,43*** 74,7±2,87*** Декабрь 36,8±1,24 22,0±0,40 2,8±0,14 8,7±0,04 10,7±0,14 44,5±0,64 137,2±0,64 0,5±0,03 0,6±0,01 58,7±1,95 72,0±3,11*** 23,4±3,11 44,7±2,90*** Примечания: С – сустентоциты, Г – гоноциты; различия с предыдущей группой достоверны: *** – при Р<0,001; ** – при Р<0,01; * – при Р<0,05. Сезонные гистохимические показатели семенника В конце сентября у самцов ондатры наблюдается хорошо заметная реакция на содержание нейтральных гликопротеинов в эпителии семенных канальцев и сульфатированных протеогликанов в белочной оболочке, интерстициальной ткани и собственной пластинке семенных канальцев и 18 канальцев rete testis. Слабая реакция отмечена на общий и катионный белки, РНК, лизин, гистидин, аргинин в эпителии извитых семенных канальцев, а также в соединительнотканных структурах семенника. На все вышеперечисленные компоненты у спермиев наблюдается интенсивная реакция. В конце ноября у самцов ондатры происходят заметные гистохимические изменения, а именно, реакция на гликоген в эпителии семенных канальцев существенно снижается. Преимущественно, гликоген обнаруживается только в отдельных канальцах, которые находятся на стадии дегенерации. Отмечается существенное снижение РНК в цитоплазме и ядрах ИЭ. Происходит снижение интенсивности реакции на нейтральные гликопротеины и сульфатированные протеогликаны в собственной пластинке канальцев, белочной оболочке, в интерстициальной ткани. Сезонная гормональная активность семенника В период спада половой активности самцов ондатры до наступления периода полового покоя помимо морфофункциональных перестроек в клетках и тканях семенника происходит и снижение уровня содержания половых гормонов в сыворотки крови. Так в начале спада половой активности самцов ондатры в последней декаде сентября концентрация тестостерона, лютропина и фоллитропина в сыворотки крови была максимальной. В течение исследуемого периода до наступления периода полового покоя в конце декабря происходит плавное снижение уровня содержания данных половых гормонов: тестостерона в 2,4 раза, лютропина в 4,5 раза и фоллитропина в 18 раз (табл. 4). Таблица 4 Концентрация половых гормонов в сыворотке крови самцов ондатры в зависимости от половой активности Месяц Тестостерон (нМ/л) ЛГ (МЕД/л) ФСГ (МЕД/л) сентябрь 68,74±1,21 0,638±0,01 0,78±0,01 октябрь 59,30±1,26*** 0,438±0,03*** 0,44±0,02*** ноябрь 49,98±0,95*** 0,276±0,01*** 0,073±0,002*** декабрь 28,82±0,64*** 0,142±0,01*** 0,044±0,002*** Примечание: различия с предыдущей группой достоверны: *** – при Р<0,001; ** – при Р<0,01; * – при Р<0,05 МОРФОФУНКЦИОНАЛЬНЫЙ ГИСТОГЕНЕЗ ПРИДАТКА СЕМЕННИКА Возрастные морфологические изменения придатка семенника Придаток семенника у самцов ондатры по строению схож с таковыми у большинства животных. Тело придатка узкое, головка его плоская, хвост придатка крупный, удлинен. Придаток семенника прикреплен к семеннику 19 очень широкой брыжейкой и в отличие от большинства млекопитающих не тесно сращен с последним, довольно подвижный. К семеннику плотно приращен головчатый конец и хвост придатка. У новорожденных щенков ондатры придаток представлен слабоизвивающимся протоком и выносящими канальцами. Эпителий выносящих канальцев простой низкостолбчатый. В 1-месячном возрасте выносящие канальцы и проток придатка становятся более извитыми. Эпителий становится простым столбчатым. Ядра эпителиоцитов округлой и округло-овальной формы, расположенные, преимущественно, в базальной части клеток. В 3 месяца эпителий становится псевдодвуслойным, появляются вставочные эпитеолиоциты. Наблюдается увеличение высоты эпителия и диаметра протока придатка. Причем увеличение вышеуказанных показателей происходит от головки к хвосту (табл. 5). В 5- месячном возрасте в эпителии протока придатка у ондатры мы выделили 4 клеточных типа: столбчатые, базальные, апикальные, бокаловидные клетки. Ядра базальных эпителиоцитов мелкие, округлой формы. Столбчатые эпителиоциты содержат реснички, проявляют признаки секреции. Происходит достоверное увеличение диаметра протока и высоты эпителия во всех областях придатка (табл. 5). В области тела протока эпителий высокостолбчатый. Ядра расположены на разных уровнях, вытянуты по длине клеток. В сторону хвоста придатка количество базальных эпителиоцитов увеличивается. Ядра некоторых эпителиоцитов придатка содержат эухроматин, что свидетельствует об их функциональной активности. У 8- месячных самцов ондатры просветы выносящих канальцев приобретают неправильную форму. Увеличивается длина стереоцилий . Ядра эпителиоцитов палочковидной формы, вытянуты по длине клетки. Свободная поверхность клеток проявляет признаки секреции. Диаметр протока и высота эпителия достоверно увеличиваются в головке, теле и недостоверно в хвосте придатка (табл. 5). У 10-месячных самцов ондатры в просвете протока придатка появляются спермии. Их больше в хвосте придатка. Нами установлено, что половая зрелость самцов ондатры, обитающих в условиях экосистемы Южного Прибайкалья, наступает в 10 месяцев. В 12-месячном возрасте изменения носят преимущественно количественный характер, который связан с некоторым увеличением высоты эпителия, диаметров протоков (табл. 5) Возрастные гистохимические показатели придатка семенника Формирование придатка семенника, дифференциация всех его структурных компонентов сопровождается заметными гистохимическими изменениями. Нами у новорожденных щенков ондатры в апикальной и базальной частях цитоплазмы некоторых эпителиоцитов выносящих Таблица 5 Морфометрические показатели придатка семенников самцов ондатры в зависимости от возраста Показатели 1 21,1±1,13 54,3±2,96*** 49,6±2,23*** 59,5±2,93*** 16,6±2,57*** 21,6±1,35*** 18,0±2,83*** 22,7±0,06*** 22,2±0,41*** 18,7±0,51*** 9,4±0,05 9,5±0,07** 9,6±0,10 82,4±5,53*** 85,0±2,75 84,7±2,23** 0,5±0,01* 0,5±0,02*** 0,5±0,01*** 3 22,0±0,60*** 116,2±5,14 91,1±5,14 120,5±5,18*** 45,21±3,75 24,7±1,67*** 53,5±2,16*** 35,5±1,00*** 36,5±0,85*** 28,2±0,94*** 9,4±0,06** 9,6±0,05 9,5±0,05*** 83,6±0,53** 84,7±2,50*** 85,4±0,60*** 0,5±0,03* 0,5±0,01 0,5±0,03*** 8 62,3±1,15*** 192,5±8,51 143,7±4,25*** 283,6±2,46*** 58,8±3,20*** 75,2±2,56*** 135,2±3,17*** 72,7±2,21*** 64,1±1,26*** 48,1±1,60*** 11,8±0,14*** 11,5±0,12*** 11,5±0,10*** 268,2±1,44*** 272,6±1,54*** 270,2±1,12*** 0,6±0,03*** 0,6±0,03*** 0,6±0,03*** 10 82,5±1,06 203,8±2,16 189,5±6,23 363,1±2,46*** 108,7±2,20 113,2±3,54 292,6±2,23*** 42,2±1,75 39,7±2,15 35,4±1,76 12,5±0,12 12,4±0,10 12,5±0,10*** 325,5±2,03 326,4±2,52 320,0±2,40*** 0,7±0,01*** 0,7±0,04** 0,7±0,01*** 12 80,0±2,50 218,6±5,78 195,6±3,54 378,7±5,72 112,6±3,14 115,2±3,47 313,8±3,05 48,6±3,82 42,5±3,56 32,8±2,12 12,6±0,12 12,4±0,07 12,6±0,12 333,1±1,87 328,3±2,93 326,8±1,53 0,7±0,03 0,7±0,01 0,7±0,03 Примечания: Г – головка придатка, Т – тело придатка, Х – хвост придатка; различия с предыдущей группой достоверны: *** – при Р<0,001; ** – при Р<0,01; * – при Р<0,05 20 Толщина капсулы (мкм) Диаметр протока Г (мкм) Т Х Диаметр просвета Г (мкм) Т Х Высота эпителия Г (мкм) Т Х Средний диаметр Г клеток (мкм) Т Х Объем ядер клеток Г (мкм3) Т Х ЯЦС (ед.) Г Т Х Новорожден 22,2±1,11 48,5±8,14 43,4±1,25 43,3±4,21 14,7±1,28 18,2±1,28 17,1±1,54 17,1±0,27*** 16,4±0,36*** 14,1±0,35*** 8,3±0,07*** 8,0±0,06*** 8,2±0,05*** 41,6±1,56*** 43,1±1,80*** 42,8±1,07* 0,5±0,01 0,5±0,02 0,5±0,01 Возраст (мес.) 5 32,6±1,00*** 126,2±5,25*** 125,2±2,53*** 192,7±6,14*** 46,8±2,14*** 56,2±2,95*** 76,4±3,72*** 49,5±0,30*** 47,1±1,24*** 39,5±0,27 9,6±0,06*** 9,6±0,06** 9,6±0,10** 83,4±1,00** 85,2±0,67 87,5±1,55*** 0,5±0,02** 0,5±0,01*** 0,5±0,02** 21 канальцев и протока придатка в незначительном количестве обнаруживается гликоген. Затем происходит его постепенное увеличение до 5-месячного возраста. В 8 месяцев содержание его не меняется, а к 10-12 месяцам происходит существенное снижение этого компонента. По нашему мнению, уменьшение гликогена связано с расходованием его для синтеза гликопротеинов для питания спермиев и для поддержания слабокислой реакции содержимого протока придатка, что является обязательным условием для нормального существования спермиев. Нейтральные гликопротеины выявляются в незначительном количестве у новорожденных щенков ондатры, затем с возрастом их количество постепенно увеличивается. Нейтральные и кислые сульфатированные гликопротеины обнаруживаются с 3-месячного возраста в цитоплазме эпителиоцитов выносящих канальцев и протока придатка семенника самцов ондатры, которые с возрастом увеличиваются. Некоторое увеличение количества кислых сульфатированных гликопротеинов, по-нашему мнению, связано с участием их в создании слабокислой среды в содержимом просвета протока придатка, которая нужна для нормальной депонации спермиев. Сиалогликопротеины выявляются в незначительном количестве в апикальной части цитоплазмы эпителиоцитов придатка у новорожденных щенков ондатры. Количество их увеличивается в 8-месяцев, что совпадает с началом полового созревания. Количество общего и катионного белков, РНК, ДНК, лизина, гистидина увеличивается в эпителии выносящих канальцев и протока придатка с возрастом. Аргинин у новорожденных и 1-месячных щенков ондатры идентифицируется в ядрах эпителиоцитов. В период обнаружения спермиев в протоке придатка семенника самцов ондатры содержание выявляемых белковых компонентов увеличивается, что дает право предположить об усилении секреторной деятельности эпителиоцитов. Гистохимическим изменениям подвергается и строма придатка. Незначительное накопление гликогена с последующим увеличением нейтральных гликопротеинов, сульфатированных протеогликанов, белковых компонентов отмечается на ранних стадиях развития. У половозрелых самцов строма придатков насыщена нейтральными гликопротеинами, общим и катионным белками, лизином, гистидином. СЕЗОННЫЕ МОРФОФУНКЦИОНАЛЬНЫЕ ИЗМЕНЕНИЯ ПРИДАТКА СЕМЕННИКА Сезонные морфологические изменения придатка семенника В конце сентября эпителий протока придатка представлен 4 клеточными типами: столбчатыми, базальными, апикальными и бокаловидными. Бокаловидные клетки встречаются преимущественно ближе 22 к хвосту придатка. Во всех частях придатка обнаруживаются спермии. Большинство ядер всех типов клеток являются активными. К концу октября у исследуемых особей происходит уменьшение диаметра протока придатка и высоты эпителия (табл. 6). Морфологически выявляются среди эпителиоцитов дегенерирующие клетки. В просвете протока спермии обнаруживаются у одного животного исследуемой группы, однако количество их значительно меньше, чем в предыдущем периоде. Таблица 6 Морфометрические показатели придатка семенника самцов ондатры в зависимости от половой активности Показатели Сентябрь Толщина капсулы (мкм) 77,6±1,33*** Диаметр протока (мкм) Г 205,7±2,23*** Т 193,1±5,47*** Х 171,0±5,67*** Диаметр просвета Г 116,7±2,57*** (мкм) Т 110,5±4,26*** Х 105,1±2,63*** Высота эпителия Г 45,5±0,95 (мкм) Т 40,5±0,83 Х 33,1±0,86** Средний диаметр Г 12,8±0,14*** клеток (мкм) Т 12,5±0,12*** Х 12,5±0,10*** Объем ядер клеток Г 330,2±1,44*** (мкм3) Т 287,6±1,60*** Х 280,2±1,10*** ЯЦС (ед.) Г 0,7±0,03*** Т 0,7±0,03** Х 0,7±0,03** Период времени (мес.) Октябрь Ноябрь 80,3±1,31*** 83,4±1,24*** 157,5±4,35*** 123,3±3,47*** 116,1±3,64*** 95,4±2,16** 104,4±5,15*** 83,2±2,42*** 73,1±2,12*** 53,1±1,56*** 51,1±3,53 45,1±3,17*** 48,1±1,60** 37,1±2,17*** 42,2±1,24*** 34,2±0,78*** 32,2±1,16** 25,2±1,26** 28,1±0,75** 23,1±1,22*** 12,5±0,12 12,6±0,12*** 12,4±0,10** 12,4±0,07 12,5±0,12*** 12,6±0,06 326,5±2,03*** 328,1±1,87*** 276,4±2,53 278,3±2,93*** 278,0±2,40** 276,8±1,53*** 0,7±0,01*** 0,6±0,03** 0,7±0,04** 0,6±0,01** 0,7±0,01*** 0,6±0,03** Декабрь 86,8±1,25 56,3±1,31 61,4±2,77 52,1±1,97 24,1±1,71 23,2±1,94 18,1±2,23 21,1±0,93 18,1±0,97 15,7±0,86 12,5±0,10 12,4±0,06 12,5±0,12 324,3±1,73 270,4±2,33 268,0±2,30 0,6±0,01 0,6±0,04 0,6±0,01 Примечания: Г – головка придатка, Т – тело придатка, Х – хвост придатка; различия с предыдущей группой достоверны: *** – при Р<0,001; ** – при Р<0,01; * – при Р<0,05. У самцов ондатры на конец ноября происходит значительное снижение всех исследуемых показателей во всех областях придатка (табл. 6). Изменение удельных объемов паренхимы, кровеносных сосудов и стромы связано с уменьшением первых. В хвосте толщина волокнисто-мышечного слоя больше, чем в головке и теле. В просвете протока придатка отмечаются скопления клеток различной формы с гиперхромными ядрами, неровно 23 очерченной оксифильной цитоплазмой. Большинство ядер клеток эпителиоцитов содержит гетерохроматин. К концу декабря у самцов ондатры отмечается достоверное существенное уменьшение морфометрических показателей протока придатка во всех его частях (табл. 6). Просвет протока придатка на всем протяжении не содержит спермии. Толщина волокнисто-мышечного слоя существенно увеличивается, во всех областях придатка, однако в хвосте она больше. Сезонные гистохимические показатели придатка семенника Сезонные гистохимические изменения связаны в основном с постепенным уменьшением количества гликогена в период с конца сентября по конец декабря в апикальной части эпителиоцитов протока придатка, клетках волокнисто-мышечного слоя и нейтральных гликопротеинов в эпителии выносящих канальцев и протока придатка, собственной пластинке, базальной мембране, волокнисто-мышечном слое. К концу декабря интенсивность реакции на белковые компоненты в цитоплазме эпителиоцитов снижается. Аргинин в последних вообще не обнаруживается, хотя ядро, собственная пластинка, адвентиция и волокнисто-мышечный слой содержат незначительное количество этого компонента. МОРФОФУНКЦИОНАЛЬНЫЙ ГИСТОГЕНЕЗ ПРЕДСТАТЕЛЬНОЙ ЖЕЛЕЗЫ Возрастные морфологические изменения предстательной железы Предстательная железа ондатры – это хорошо выраженное железистое образование, которое лежит в тазовой полости дорсально. Железа расположена на уровне второго-третьего крестцовых позвонков. У новорожденных и 1-месячных щенков ондатры предстательная железа представлена немногочисленными эпителиальными тяжами, отходящими от эпителия уретрального канала. Формирование железистых компонентов простаты происходит путем их последующего ветвления. Концевые отделы представлены скоплением недифференцированных эпителиоцитов. В крупных выводных протоках эпителий состоит из 2-3 слоев эпителиоцитов. Ближе к каналу уретры, где видны просветы, эпителий псевдомногослойный. При качественной окраске на функциональную активность ядер эпителиоцитов обнаружено подавляющее большинство клеток содержат гетерохроматин, что свидетельствует о низкой функциональной активности органа. Дальнейший рост простаты в 3-5 месячном возрасте характеризуется разрастанием железистых структур по типу сложноветвящейся железы. Отмечается разделение крупных выводных протоков на более мелкие, которые в дальнейшем, продолжая делиться, формируют секретирующие отделы. По-видимому, процесс дифференцировки эпителия из многослойного недифференцированного в однослойный сопровождается 24 слущиванием клеток поверхностных слоев эпителиального пласта. Происходит увеличение числа тканевых базофилов в строме железы, что свидетельствует о продолжающемся росте и развитие органа (табл. 7). Простата 8-месячных самцов ондатры характеризуется преобладанием железистой паренхимы и кровеносных сосудов над соединительнотканной стромой. Паренхима органа представлена секретирующими отделами и выводными протоками, которые имеют разное строение и размеры. Для мелких выводных протоков характерно наличие простого столбчатого эпителия. Более крупные, возле канала уретры выстланы псевдомногослойным эпителием. Увеличивается количество тканевых базофилов (табл. 7). К 10-12-месячному возрасту предстательная железа самцов ондатры является хорошо выраженным дольчатым образованием, что совпадает с периодом обнаружения спермиев в просвете протока придатка, свидетельствующих о наступлении половой зрелости. Эпителий секретирующих отделов представлен столбчатыми гландулоцитами с мелкими округло-овальной формы гиперхромными ядрами, расположенными в зависимости от наличия секрета на разных уровнях. В некоторых участках наблюдается разрушение апикальной части гландулоцитов, содержимое которой выделяется в просвет отделов. Особенности секреторного процесса гландулоцитов позволяет нам судить о наличии апокринного типа секреции. Возрастные гистохимические показатели предстательной железы В предстательной железе новорожденных и 1-месячных щенков ондатры в цитоплазме гландулоцитов гликоген идентифицируется в небольших количествах. Отмечается его неодинаковое распределение в структурах железы. Клетки поверхностного слоя содержат больше этого компонента, чем клетки, расположенные ближе к базальной мембране, в цитоплазме соединительнотканных клеток в волокнах уретральной мышцы. В недифференцированных клеточных скоплениях реакция на гликоген менее интенсивная. Это дает право судить, что процесс дифференцировки более активно протекает в дистальных отделах железистых образований. С первых дней после рождения щенков ондатры идентифицируются нейтральные и сульфатированные гликопротеины, которые локализуются в эпителии крупных выводных протоков и в просвете. С возрастом количество их увеличивается, а в железе 1- месячных щенков они находятся не только в эпителии выводных протоков, но и в формирующихся ацинусах. Это, по-нашему мнению, указывает на то, что процессы морфологической и гистохимической дифференцировки берут начало у места отхождения эпителиальных тяжей от эпителия уретрального канала и идут по направлению к дистальным отделам. Таблица 7 Морфометрические показатели предстательной железы самцов ондатры в зависимости от возраста Показатели 1 70,7±9,00*** 25,0±1,60** 3 75,7±6,50 32,0±1,20*** 19,4±2,05 19,4±1,00** 8,1±0,15 8,1±0,14 36,0±5,04 36,8±4,38 0,5±0,01 0,5±0,02 20,0±3,23 23,4±2,27 8,2±0,10 8,1±0,12 36,3±5,08*** 37,0±4,62*** 0,5±0,01* 0,5±0,02* 23,0±1,46 21,0±3,26 8,2±0,13** 8,1±0,10** 38,0±4,02 38,2±5,40 0,5±0,03 0,5±0,01 22,6±4,24** 19,4±2,74** 8,3±0,08 8,3±0,06 38,5±4,27*** 38,6±4,62*** 0,5±0,02 0,5±0,01 8 69,0±2,14*** 42,4±4,50*** 10 42,2±2,14*** 47,8±7,12*** 12 40,5±2,04 55,6±10,72 28,7±0,65** 27,0±3,30** 8,3±0,06** 8,3±0,05** 40,1±5,00** 40,7±5,40** 0,5±0,03** 0,5±0,03** 21,0±2,42 19,5±0,73 8,5±0,12 8,4±0,14 42,1±1,31*** 40,2±4,60** 0,6±0,01 0,6±0,04 22,1±1,94 20,0±2,12 8,5±0,15 8,4±0,14 43,0±2,63 41,0±3,24 0,6±0,03 0,6±0,01 Примечания: КО – концевые отделы, ВП – выводные протоки; различия с предыдущей группой достоверны: *** – при Р<0,001; ** – при Р<0,01; * – при Р<0,05 25 Толщина капсулы (мкм) Кол-во тканевых базофилов Высота эпителия КО (мкм) ВП Средний диаметр КО клеток (мкм) ВП Объем ядер клеток КО (мкм3) ВП ЯЦС (ед.) КО ВП Новорожден. 113,6±1,00** 23,0±2,80*** Возраст (мес.) 5 77,1±5,56*** 37,0±0,80*** 26 В 3 месяца в железистых структурах наблюдается дальнейшее увеличение содержания нейтральных, сульфатированных гликопротеинов и уменьшение гликогена. С 5 месяцев в апикальных зонах некоторых эпителиоцитов концевых отделов и выводных протоков, в секрете обнаруживаются сиалогликопротеины и кислые сульфатированные гликопротеины значительно у половозрелых самцов ондатры. Это позволяет предположить о начале секреторной деятельности железы и считать концевые отделы и выводные протоки секретирующими структурами. Количество нейтральных и сульфатированных гликопротеинов существенно повышается в 10-12 месяцев. Они, как правило, локализуются в цитоплазме отдельных гландулоцитов секретирующих отделов и эпителиоцитов выводных протоков, в содержимом просвета. Как видно из приведенных данных, становление гистохимической специализации гландулоцитов происходит ступенчато с накопления гликогена, появлением сначала нейтральных, а после кислых сульфатированных гликопротеинов и сиалогликопротеинов. После рождения щенков ондатры белковые компоненты обнаруживаются в эпителии простаты, соединительной ткани. Происходит возрастное увеличение белковых компонентов. Их несколько больше в апикальной части клеток. В цитоплазме эпителиоцитов выводных протоков и гландулоцитов секретирующих отделов с 5 месяцев отмечается незначительное содержание аргинина, однако их ядра весьма отчетливо реагируют на этот компонент. Морфологических различий в специализации клеток эпителия простаты (подразделение их на мукоциты и сероциты) нами не отмечено. Вероятно один и тот же клеточный тип в простате способен вырабатывать одновременно или поочередно различные вещества: либо слизь, либо белковые зернышки. Анализируя полученные нами гистохимичекие показатели, можно сказать, что эпителий простаты проявляет значительную гистохимическую гетерогенность и секретирует углеводно-белковые компоненты. Однако обращает на себя внимание относительно высокое содержание в составе секрета предстательной железы ондатры углеводных компонентов, в частности сиалогликопротеинов и кислых сульфатированных гликопротеинов. СЕЗОННЫЕ МОРФОФУНКЦИОНАЛЬНЫЕ ИЗМЕНЕНИЯ ПРЕДСТАТЕЛЬНОЙ ЖЕЛЕЗЫ Сезонные морфологические изменения предстательной железы К концу сентября предстательная железа у самцов ондатры является хорошо выраженным дольчатым образованием. Паренхима преобладает над стромой . Выводные протоки выстланы однослойным столбчатым эпителием. Эпителий выводных протоков секретирующий. Секретирующие отделы в долях окружены тонкими соединительнотканными прослойками. 27 Практически все ядра клеток секретирующих отделов и выводных протоков являются функционально активными. Сезонные морфологические перестройки предстательной железы самцов ондатры характеризуются постепенным увеличением удельного объема стромы и, соответственно уменьшением удельных объемов паренхимы и кровеносных сосудов. Отмечается утолщение соединительнотканных перегородок. Уменьшение высоты эпителия концевых отделов и выводных протоков незначительно. Таблица 8 Морфометрические показатели предстательной железы самцов ондатры в зависимости от половой активности Показатели Толщина капсулы (мкм) Кол-во тканевых базофилов Высота КО эпителия (мкм) ВП Средний КО диаметр клеток ВП (мкм) Объем ядер КО 3) клеток (мкм ВП ЯЦС (ед.) КО ВП Сентябрь 44,8±2,96*** Период времени (мес.) Октябрь Ноябрь 42,4±1,28*** 48,4±3,92*** Декабрь 52,4±1,28 51,6±2,48*** 47,6±2,72*** 37,6±4,72*** 32,8±1,76 25,1±2,96 22,7±1,27 8,5±0,15 8,4±0,14 22,7±3,38 20,1±2,55 8,5±0,12*** 8,4±0,14 23,7±5,04 22,0±3,34 8,3±0,06 8,3±0,05 20,7±5,26 19,6±3,76 8,3±0,08 8,3±0,06 43,0±2,63*** 41,0±3,24 0,6±0,03 0,6±0,01 42,1±1,31*** 40,2±4,60 0,6±0,01 0,6±0,04 40,1±5,00** 40,7±5,40** 0,5±0,03 0,5±0,03 38,5±4,27 38,6±4,62 0,5±0,02 0,5±0,01 Примечания: КО – концевые отделы, ВП – выводные протоки; различия с предыдущей группой достоверны: *** – при Р<0,001; ** – при Р<0,01; * – при Р<0,05 Сезонные гистохимические показатели предстательной железы Сезонные гистохимические изменения в простате ондатры связаны с постепенным уменьшением гликогена в соединительнотканной капсуле, межальвеолярной соединительной ткани, эпителии секретирующих отделов. В состоянии относительного полового покоя уменьшается содержание интенсивности реакции на нейтральные гликопротеины и повышается на сульфатированные гликопротеины и протеогликаны. Сиалогликопротеины не выявляются. Незначительно происходит уменьшение интенсивности реакции на катионный белок, РНК и гистидин. Аргинин в цитоплазме гландулоцитов не выявляется. Несомненно, биологические особенности воспроизводительной функции самцов ондатры можно увидеть в специфичности секреции предстательной железы и ее зависимости от времени года. 28 МОРФОФУНКЦИОНАЛЬНЫЙ ГИСТОГЕНЕЗ ПУЗЫРЬКОВИДНОЙ ЖЕЛЕЗЫ Возрастные морфологические изменения пузырьковидной железы Пузырьковидная железа ондатры – парное дольчатое образование, располагающееся дорсально от мочевого пузыря и латерально от железистой части семяпровода. Пузырьковидная железа у новорожденных щенков ондатры представляет собой хорошо выраженное дольчатое образование и разделена на доли широкими прослойками соединительной ткани. В каждой доле заметно разделение их на мелкие дольки, состоящие из железистых трубочек и эпителиальных тяжей. Помимо железистых трубочек в железе обнаруживаются канальцы мелкого размера с округлым просветом и крупные с просветом неправильной формы. Железистые трубочки выстланы псевдомногослойным эпителием. Мы выделили два типа эпителиоцитов: базальные и столбчатые. Базальные эпителиоциты с округлыми ядрами, а столбчатые с ядрами округло-овальной формы, расположенными в апикальной части клеток. Встречаются столбчатые эпителиоциты с палочковидными формами клеток. У 1-3-месячных самцов ондатры продолжаются процессы ветвления более крупных железистых трубочек на более мелкие. Происходит увеличение высоты эпителия концевых отделов и выводных протоков пузырьковидной железы (табл. 9). Характерной особенностью этих возрастов является выявление сульфатированных гликопротеинов в апикальной зоне клеток отдельных гландулоцитов и в просвете трубочек. Незначительно усиливается реакция на сульфатированные протеогликаны в соединительнотканных волокнах и стенках сосудов. В 5 месяцев железа представлена большим количеством долей, каждая из которых окружена волокнистой соединительнотканной капсулой с подлежащим мышечным слоем, имеющим пучки гладкомышечных клеток, расположенных продольно и циркулярно. Цитоплазма столбчатых эпителиоцитов окрашивается оксифильно. Ядра округло – овальной формы находятся в апикальной зоне клеток. Ядра базальных эпителиоцитов или крупные и светлые с мелкими глыбками хроматина, или мелкие, неправильной формы, гиперхромные. Высота эпителия изменяется незначительно (табл. 9). У самцов ондатры 8-10-12 месяцев паренхима органа представлена многочисленными железистыми трубочками разных размеров, которые окружены тонкими плотными прослойками соединительной ткани. В отдельных участках секрет из гландулоцитов выделяется в просвет без видимого нарушения апикальной части клеток, поэтому мы считаем, что пузырьковидная железа ондатры характеризуется наличием мерокринового типа секреции. Возрастные морфометрические показатели сопровождаются постепенным увеличением удельных объемов паренхимы и кровеносных Таблица 9 Морфометрические показатели пузырьковидной железы самцов ондатры в зависимости от возраста Показатели 1 73,8±8,65*** 13,8±2,24* 3 71,0±3,24 12,6±1,52*** 22,6±0,68 23,2±3,78** 8,1±0,15 8,1±0,14 36,0±5,04 36,8±4,38 0,5±0,01 0,5±0,02 24,5±0,96 28,1±2,16 8,2±0,10 8,1±0,12 36,3±5,08*** 37,0±4,62*** 0,5±0,01* 0,5±0,02* 24,0±2,53 25,6±6,31** 8,2±0,13** 8,1±0,10** 38,0±4,02 38,2±5,40 0,5±0,03 0,5±0,01 26,5±2,24** 32,1±4,25 8,3±0,08 8,3±0,06 38,5±4,27*** 38,6±4,62*** 0,5±0,02 0,5±0,01 8 53,5±8,14*** 20,2±3,84*** 10 45,5±2,42*** 37,4±5,84*** 12 43,7±1,65 41,4±4,88 32,2±3,74** 30,5±6,52** 8,3±0,06** 8,3±0,05** 40,1±5,00** 40,7±5,40** 0,5±0,03** 0,5±0,03** 23,4±1,93 24,1±5,18 8,5±0,12 8,4±0,14 42,1±1,31*** 40,2±4,60** 0,6±0,01 0,6±0,04 22,1±1,46 21,5±3,22 8,5±0,15 8,4±0,14 43,0±2,63 41,0±3,24 0,6±0,03 0,6±0,01 Примечания: ВП – выводные протоки, КО – концевые отделы; различия с предыдущей группой достоверны: *** – при Р<0,001; ** – при Р<0,01; * – при Р<0,05 29 Толщина капсулы (мкм) Кол-во тканевых базофилов Высота эпителия КО (мкм) ВП Средний диаметр КО клеток (мкм) ВП Объем ядер клеток КО (мкм3) ВП ЯЦС (ед.) КО ВП Новорожден. 117,0±0,92** 16,4±1,00*** Возраст (мес.) 5 71,0±8,44*** 14,0±3,20*** 30 сосудов с соответствующим уменьшением удельного объема стромы, а так же незначительным увеличением высоты эпителия концевых отделов и выводных протоков железы (табл. 9). Возрастные гистохимические показатели пузырьковидной железы Гистохимичский анализ, проведенный у новорожденных щенков ондатры, свидетельствует о присутствии существенного количества гликогена в цитоплазме гландулоцитов формирующихся железистых структур. Его меньше в клетках соединительной ткани и стенках кровеносных сосудов. У 1-месячных щенков ондатры значительное количество этого компонента содержится в просвете железистых трубочек, а в соединительнотканных и гладкомышечных клетках содержание его снижается. В 10-12-месячном возрасте отмечается снижение интенсивности реакции на гликоген. Наличие большого количества гликогена и существенное снижение его в эпителии пузырьковидных желез ондатры в период полового созревания является видовой особенностью и, по нашему мнению, гликоген расходуется на снабжение спермиев энергией для движения. С первых дней после рождения щенков ондатры в пузырьковидной железе отмечается наличие нейтральных гликопротеинов в собственной пластинке, а сульфатированных протеогликанов в соединительно-тканной капсуле. В эпителии и в содержимом просвета железистых трубочек в 1месячном возрасте начинают выявляться нейтральные и сульфатированные гликопротеины. Они выявляются, преимущественно, в апикальной зоне клеток. С возрастом количество этих компонентов возрастает. Характерной особенностью является наличие у самцов ондатры в 8 месяцев кислых сульфатированных гликопротеинов и сиалогликопротеинов. Содержание сульфатированных протеогликанов и нейтральных гликопротеинов в соединительной ткани железы с возрастом уменьшается. Количество общего белка, РНК, лизина, гистидина в апикальной части цитоплазмы, а ДНК и аргинина в ядрах гландулоцитов, с возрастом увеличивается. Начинают обнаруживаться вышеперечисленные белковые компоненты в цитоплазме и ядрах гландулоцинов с 5-месячного возраста. Свободный катионный белок идентифицируется с 1-месячного возраста в ядрах эпителиоцитов, а в 5 месяцев в цитоплазме эпителиоцитов с последующим увеличением его до 8-месячного возраста. В 10-12 месяцев его содержание не изменяется, по сравнению с предыдущим сроком. СЕЗОННЫЕ МОРФОФУНКЦИОНАЛЬНЫЕ ИЗМЕНЕНИЯ ПУЗЫРЬКОВИДНОЙ ЖЕЛЕЗЫ Сезонные морфологические изменения пузырьковидной железы В конце сентября паренхима пузырьковидной железы состоит из многочисленных железистых трубочек, которые отличаются своими 31 размерами друг от друга. Железистые трубочки окружены тонкими прослойками соединительной ткани. Эпителий железистых трубочек высокостолбчатый с единичными базальными эпителиоцитами. Цитоплазма столбчатых эпителиоцитов окрашена оксифильно, а базальных эпителиоцитов слабооксифильно. Ядра столбчатых эпителиоцитов округлоовальной формы, расположенные в апикальной части клеток. В конце декабря в пузырьковидной железе отмечается утолщение соединительнотканных перегородок и, как следствие, уменьшение величины секретирующих отделов. Эпителий секретирующих отделов представлен столбчатыми гландулоцитами с округлой формы ядрами, расположенными на разных уровнях. Выводные протоки выстланы низкостолбчатым эпителием. Высота эпителия концевых отделов и выводных протоков незначительно уменьшается. Существенно снижается количество тканевых базофилов в соединительнотканных прослойках железы (табл. 10). Таблица 10 Морфометрические показатели пузырьковидной железы самцов ондатры в зависимости от половой активности Показатели Толщина капсулы (мкм) Кол-во тканевых базофилов Высота КО эпителия (мкм) ВП Средний КО диаметр клеток ВП (мкм) Объем ядер КО 3) клеток (мкм ВП ЯЦС (ед.) КО ВП Сентябрь 44,6±4,64*** Период времени (мес.) Октябрь Ноябрь 42,6±2,88*** 46,6±4,72*** Декабрь 53,5±8,14 45,0±2,80*** 43,0±2,40*** 34,4±3,28*** 25,8±5,84 30,5±4,27 32,1±1,83 8,5±0,15 8,4±0,14 25,8±5,25 24,1±2,68 8,5±0,12 8,4±0,14 25,4±6,21 23,0±4,32 8,3±0,06 8,3±0,05 23,5±5,15 19,7±3,27 8,3±0,08 8,3±0,06 43,0±2,63*** 41,0±3,24** 0,6±0,03 0,6±0,01 42,1±1,31*** 40,2±4,60** 0,6±0,01 0,6±0,04 40,1±5,00** 40,7±5,40** 0,5±0,03 0,5±0,03 38,5±4,27 38,6±4,62 0,5±0,02 0,5±0,01 Примечания: КО – концевые отделы, ВП – выводные протоки; различия с предыдущей группой достоверны: *** – при Р<0,001; ** – при Р<0,01; * – при Р<0,05 Сезонные гистохимические показатели пузырьковидной железы Гистохимический анализ проведенный в структурных элементах пузырьковидной железы самцов ондатры в конце сентября показал, что гликоген в незначительном количестве в виде мелких зерен и гранул выявляется в просвете железистых трубочек. В клетках соединительной ткани его несколько больше. 32 Реакция на нейтральные и сульфатированные гликопротеины в гландулоцитах и содержимом просвета довольно интенсивная. Незначительное содержание общего и свободного катионного белков, РНК, лизина, гистидина отмечается в апикальной зоне клеток цитоплазмы гландулоцитов. Существенное содержание белковых компонентов наблюдается в соединительнотканных структурах и гладкомышечных клетках. В конце декабря незначительное количество гликогена содержится в соединительнотканной капсуле, межальвеолярной соединительной ткани. Интенсивность реакции на нейтральные гликопротеины существенно ослабла в цитоплазме гландулоцитов и эпителиоцитов выводных протоков, в строме. Реакция на сульфатированные гликопротеины заметная в цитоплазме эпителиоцитов выводных протоков, а в содержимом просвета и в секретирующих отделах их очень мало. Довольно интенсивная реакция на содержание сульфатированных протеогликанов в соединительной ткани. Незначительное увеличение общего белка, лизина, гистидина отмечено также в соединительнотканных структурах и гладкомышечных клетках. МОРФОФУНКЦИОНАЛЬНЫЙ ГИСТОГЕНЕЗ ЩИТОВИДНОЙ ЖЕЛЕЗЫ Возрастные морфологические изменения щитовидной железы У щитовидной железы самцов ондатры форма долей является видовой особенностью. Для нее характерна трапециевидная форма. Доли железы соединяются тонким паренхиматозным перешейком. При исследовании гистоструктуры щитовидной железы самцов ондатры нами было установлено, что она принадлежит к смешанно-фолликулярному типу строения с преобладанием мелких фолликулов. К моменту рождения у самцов ондатры щитовидная железа имеет дефинитивное строение, то есть железа морфологически сформирована и проявляет наименьшую функциональную активность по сравнению с остальными возрастными периодами. Преобладание ядер тиреоцитов содержащих гетерохроматин свидетельствует о низкой функциональной активности органа на момент рождения. У 1-месячных самцов ондатры мы отметили повышение почти в 2 раза функциональной активности щитовидной железы, по сравнению с новорожденными. Начиная с 1-месячного возраста постнатального периода развития, происходит плавное увеличение всех исследуемых показателей до 3-месяцев (табл. 11). В 5-месяцев постнатального периода развития происходит заметная активизация функции щитовидной железы, о чем свидетельствует уменьшение толщины капсулы органа, увеличение остальных функционально значимых морфологических показателей и существенное снижение индекса Брауна (табл. 11). Подавляющее большинство клеток тиреоидного эпителия и что характерно коллоид содержат эухроматин. 33 Пик активности щитовидной железы самцов ондатры отмечен в 8месячном возрасте. У самцов ондатры обитающих в условиях экосистемы Южного Прибайкалья это совпадает с началом полового созревания. Почти в два раза происходит уменьшение толщины капсулы органа, существенно, по сравнению с предыдущим возрастом увеличиваются остальные микрометрические характеристики железы. Заметно снижается индекс Брауна (табл. 11). К 10-месячному возрасту, толщина капсулы становится минимальной по сравнению с остальными возрастами, что свидетельствует о щитовидной железе ондатры в этом периоде как о сформированном в функциональном отношении органе. Однако функциональная активность железы в этом возрасте ослабевает, о чем свидетельствует снижение числа фолликулов и значительное увеличение индекса Брауна. В этот период у самцов ондатры наступает половое созревание и снижение тиреоидной активности железы по-видимому связано с подготовкой организма животных для размножения. Снижение функциональной активности железы способствует уменьшению потерь энергии и сохранению жизненных сил для активного сперматогенеза и в связи с повышением половой активности самцов (табл. 11). В 12 месяцев функциональная активность щитовидной железы продолжает снижаться. В этом возрасте показатель индекса Брауна самый высокий по сравнению с остальными возрастными периодами постнатального развития самцов ондатры. Снижение функции железы в этом возрасте связано с взрослением животных. Возрастные гистохимические показатели щитовидной железы У новорожденных щенков ондатры отмечено отсутствие запасов гликогена в щитовидной железе, слабая реакция на ДНК и отсутствие аргинина в ядрах тиреоцитов. Общий белок в значительном количестве содержится в наружной капсуле органа, что позволяет нам право предположить, что в этом возрасте идет формирование щитовидной железы как органа, так как известно, что белок в живом организме используется как строительный материал. В 1-месячном возрасте обнаруживается в больших количествах гликоген, который расходуется до 3-месячного возраста и увеличивается количество других исследуемых углеводных компонентов. Отсутствие гликогена в 3месячном возрасте и большое содержание гликопротеинов в коллоиде железы мы связываем с использованием его как энергетического и исходного материала для синтеза более сложных углеводосодержащихсоединений, необходимых для развития и функции органа. Наличие в ядрах тиреоцитов ДНК и увеличении РНК в цитоплазме тиреоцитов свидетельствует о начале активации гормоносинтезирующей функции железы. В 5 месяцев постнатального периода развития содержание общего белка увеличилось в коллоиде железы, в цитоплазме и ядрах тиреоцитов Таблица 11 Морфометрические показатели щитовидной железы самцов ондатры в зависимости от возраста Показатели 1 39,7±0,20*** 3 33,7±0,70*** 57,2±2,30*** 70,4±0,60* 84,8±7,40*** 116,2±5,00*** 155,4±3,20*** 61,4±3,10 62,8±2,80 27,3±1,50*** 40,5±0,06** 40,9±0,20*** 42,1±0,10*** 38,2±0,20*** 54,0±0,70*** 70,5±0,50 121,7±3,80*** 697,9±3,00*** 778,0±12,20*** 733,1±4,30*** 528,9±7,60*** 828,7±23,30*** 1457,5±18,80 5,3±0,02*** 10,4±0,06*** 13,4±0,05*** 15,0±0,04 15,0±0,02* 15,2±0,10*** 10,8±0,04 29,5±0,19*** 117,3±0,93*** 201,6±0,36*** 241,5±0,65 240,0±0,60*** 243,9±0,63*** 119,0±0,22 28,4±0,75*** 0,5±0,01*** 3,12 49,4 ± 3,28 83,4±1,84*** 150,7±4,30 0,3±0,01* 0,2±0,01*** 1,87 1,06 46,0 ± 2,00*** 34,8±1,04*** 159,4±3,97*** 0,2±0,05*** 0,80 9,4±1,52 201,3±4,42 0,2±0,03 0,54 5,8±1,44 208,4±4,76*** 0,2±0,03*** 1,54 отсутствуют 84,7±1,48 0,2±0,05 4,52 отсутствуют 10 9,7±0,10 12 9,4±0,20 Примечание: различия с предыдущей группой достоверны: *** – при Р<0,001; ** – при Р<0,01; * – при Р<0,05 34 Толщина капсулы (мкм) Число фолликулов в поле зрения Средний диаметр (фолликулов (мкм) Средний объем фолликулов (мкм3) Высота тиреоцитов (мкм) Средняя площадь тиреоцитов (мкм2) Объем ядер (мкм3) ЯЦС (ед.) Индекс Брауна (ед) Кол-во тканевых базофилов Н/Р 44,6±0,24*** Возраст (мес.) 5 8 28,4±0,10*** 16,3±0,20*** 35 осталось без изменений, а в наружной капсуле уменьшилось. Увеличение содержания общего белка в коллоиде по-нашему мнению связано усилением гормонообразования и активного поступления гормона из коллоида в кровь. Это также подтверждается и усилением качественной реакции на ДНК в ядрах тиреоцитов и яркой пиронинофилией в их цитоплазме, свидетельствующей об увеличении количества РНК, что свидетельствует об активации белкового синтеза, когда усиливаются процессы пролиферации. В 8 месяцев гистохимические изменения характеризуются увеличением количества содержания кислых сульфатированных протеогликанов в соединительнотканных прослойках железы. Усиливается реакция на гиалурунаты при сохранении характера их распределения по сравнению с предыдущим возрастом. Увеличилось содержание общего белка в коллоиде железы, а в цитоплазме тиреоцитов и наружной капсуле уменьшилось. Отмечается снижение уровня содержания РНК и ДНК в структурных элементах органа. В 10-12 месячном возрасте происходит уменьшение количества содержания кислых сульфатированных протеогликанов в соединительнотканных прослойках железы. В цитоплазме тиреоцитов снижается интенсивность реакции на кислые сульфатированные гликопротеины, общий белок, РНК. Уменьшается содержание ДНК в ядрах тиреоцитов. Возрастная гормональная активность щитовидной железы Функция гормональной активности щитовидной железы самцов ондатры зависела от возраста животных. Так, в период раннего онтогенеза у 1-месячных щенков была установлена наиболее высокая концентрация трийодтиронина и тироксина в сыворотке крови (табл. 12). Начиная с 3-месячного возраста концентрация тиреоидных гормонов начинает снижаться. Таблица 12 Концентрация тиреоидных гормонов в сыворотке крови самцов ондатры в зависимости от возраста Возраст Т3 Т3св Т4 Т4св (мес) (нМ/л) (пМ/л) (нМ/л) (пМ/л) н/р 1,3±0,01 3,0±0,50 18,3±0,62 7,2±0,02 1 3,8±0,13*** 4,8±0,20*** 56,0±3,94*** 9,7±0,65 3 2,9±0,20*** 3,0±0,07*** 54,6±3,87 8,1±0,38 5 2,0±0,06*** 2,6±0,18 32,3±0,64*** 6,6±0,33 8 2,0±0,01 2,3±0,20 21,5±0,58*** 5,0±0,73 10 3,7±0,18*** 4,7±0,23*** 38,4±0,74*** 9,6±0,53 12 3,7±0,14 5,0±0,34 44,0±0,86*** 9,7±0,60 Примечание: различия с предыдущей группой достоверны: *** – при Р<0,001; ** – при Р<0,01; * – при Р<0,05 36 В 8 месяцев постнатального периода развития у самцов ондатры была установлена наименьшая концентрация тиреоидных гормонов в сыворотки крови. Однако в 10 - 12 месячном периоде постнатального онтогенеза содержание тиреоидных гормонов вновь возрастает, что совпадает с половым созреванием самцов ондатры в данном регионе. СЕЗОННЫЕ МОРФОФУНКЦИОНАЛЬНЫЕ ИЗМЕНЕНИЯ ЩИТОВИДНОЙ ЖЕЛЕЗЫ Сезонные морфологические изменения щитовидной железы В период начала спада половой активности у самцов ондатры щитовидная железа является активно функционирующим органом. Об активной функции щитовидной железы на данном этапе спада половой активности ондатры свидетельствует низкий показатель индекса Брауна (табл. 13), который самый низкий в периоде от начала спада до наступления периода полового покоя. К концу октября во время спада половой активности у ондатры в зоне Южного Прибайкалья, происходит плавное снижение исследуемых микроморфометрических показателей. Однако средняя площадь тиреоцитов существенно преобладает по сравнению с предыдущим периодом. Повидимому, это связано с преобладанием в паренхиме железы фолликулов среднего и крупного размеров. Индекс Брауна по-прежнему имеет невысокий показатель, что свидетельствует об активности щитовидной железы в данное время года (табл. 13). При оценке функциональной активности ядер тиреоцитов по методу Яцковского выявлено, что в щитовидной железе преобладают тиреоциты с синей окраской ядер, но наряду с этим, встречаются как красные неактивные клетки, содержащие гетерохроматин, так и с промежуточной окраской. В конце ноября значительно увеличивается индекс Брауна, диаметр и средний объем фолликулов. Количество фолликулов в поле зрения почти в два раза снижается по сравнению с предыдущими периодами. Средняя площадь тиреоцитов также имеет тенденцию к уменьшению. При оценке функциональной активности ядер тиреоцитов отмечается снижение количества ядер содержащих эухроматин. Все это свидетельствует о начале спада функциональной активности щитовидной железы. Существенное снижение всех исследуемых показателей щитовидной железы у самцов ондатры в декабре можно считать адаптивной реакцией организма для уменьшения затрат энергии. Этот период характеризуется самым высоким индексом Брауна, максимальным диаметром и объемом фолликулов, а также наименьшим их числом в поле зрения (табл. 13). В этот же период у самцов ондатры в этой местности наступает период полового покоя. Понижение тиреоидной активности способствует уменьшению потерь энергии и увеличению жизнестойкости во время зимней бескормицы. 37 Таблица 13 Морфометрические показатели щитовидной железы самцов ондатры в зависимости от половой активности Показатели Толщина капсулы (мкм) Число фолликулов в поле зрения Средний диаметр фолликулов (мкм) Средний объем фолликулов (мкм3) Высота тиреоцитов (мкм) Средняя площадь тиреоцитов (мкм2) Объем ядер (мкм3 ) ЯЦС (ед.) Индекс Брауна (ед.) Период времени (мес.) Октябрь Ноябрь 8,0±0,10*** 9,3±0,10*** 124,6±3,70* 66,2±3,20*** Декабрь 9,6±0,10 68,6±3,20 36,2±0,20*** 37,5±0,30*** 71,0±0,50 520,6±8,40*** 532,3±11,20 14,7±0,10*** 14,3±0,03*** 14,1±0,07* 10,9±0,05 120,3±0,84*** 230,4±0,81*** 212,1±0,62*** 205,0±0,60 206,6±4,58 0,2±0,05 0,5 199,3±1,36*** 0,3±0,03*** 0,6 156,4±3,32*** 0,2±0,03*** 1,8 83,0±1,74 0,2±0,04 4,5 Сентябрь 6,2±0,20** 107,0±6,80 54,0±0,50*** 837,1±24,70*** 1461,5±18,37 Примечание: различия с предыдущей группой достоверны: *** – при Р<0,001; ** – при Р<0,01; * – при Р<0,05. Сезонные гистохимические показатели щитовидной железы В гистохимическом выражении в период спада половой активности самцов ондатры в структурных элементах щитовидной железы отмечается снижение уровня содержания нейтральных гликопротеинов. В отношении белковых компонентов выявлено снижение интенсивности реакции на общий белок в цитоплазме и ядрах тиреоцитов, а также в коллоиде железы, незначительное содержание РНК в цитоплазме тиреоцитов и ДНК в ядрах. Сезонная гормональная активность щитовидной железы Проведенные нами исследования выявили зависимость гормональной активности щитовидной железы самцов ондатры от сезонных изменений окружающей среды. Так, например, в конце сентября, когда в данном регионе, как правило, отмечается теплая погода уровень всех исследуемых гормональных показателей щитовидной железы самый высокий по сравнению с остальными месяцами. В дальнейшем, в конце октября, происходит снижение уровня тиреоидных гормонов. Это связано с изменением погодных условий в сторону низких температур, а также с началом наступления у самцов ондатры, обитающих в данном регионе стадии полового покоя. С наступлением холодов в конце ноября, продолжается спад функциональной активности щитовидной железы. 38 Зимнее снижение тиреоидной активности в конце декабря, обусловлено адаптивной реакцией организма для уменьшения затрат и энергии. Из приведенных данных (табл. 13) видно, что наименьшие показатели уровня содержания тиреоидных гормонов в сыворотке крови отмечены в конце декабря. Таблица 14 Концентрация тиреоидных гормонов в сыворотке крови самцов ондатры в зависимости от половой активности Месяц Т3(нМ/л) Т3св(пМ/л) Т4(нМ/л) Т4св(пМ/л) сентябрь 3,6±0,13 4,6±0,18 43,6±0,67 9,7±0,37 октябрь 2,3±0,07*** 3,3±0,20*** 37,8±0,63*** 8,2±0,38 ноябрь 2,1±0,02** 2,6±0,03** 22,6±0,90*** 6,6±0,32 декабрь 1,8±0,11** 2,2±0,03*** 18,5±0,27*** 5,3±0,22 Примечание: различия с предыдущей группой достоверны: *** – при Р<0,001; ** – при Р<0,01; * – Р<0,05. МОРФОФУНКЦИОНАЛЬНЫЙ ГИСТОГЕНЕЗ НАДПОЧЕЧНИКА Возрастные морфологические изменения надпочечника У новорожденных самцов ондатры надпоченик покрыт соединительнотканной капсулой, от которой внутрь паренхимы органа отходит тонкая сеть коллагеновых волокон, поддерживающая секреторные клетки. В гистоструктуре надпочечника отмечена слабая дифференцировка коркового и мозгового вещества. Мозговое вещество занимает большую часть органа. Кора надпочечника характеризуется наличием клубочковой и пучковой зон и отсутствием сетчатой зоны. В 1-месячном возрасте в гистоструктуре надпочечника самцов ондатры происходит дифференциация функциональных зон. Появляется сетчатая зона, границы между зонами становятся четкими. Мозговое вещество занимает большую часть органа. В коре надпочечника сетчатая и клубочковая зоны занимают примерно равный между собой объем, а пучковая зона самая большая (табл. 15). Удельная доля мозгового вещества превалирует над корковым в поснатальном периоде онтогенеза в структуре надпочечника до 3-месячного возраста (табл. 15), после происходит ее уменьшение в пользу коркового вещества и достижения минимального значения в 10 месяцев (табл. 15). В корковом веществе надпочечника объем пучковой зоны на всем протяжении постнатального периода является доминирующим по отношению к остальным функциональным зонам. Наибольшего своего значения она достигает в 3 месяца (табл. 15), что совпадает с периодом роста и развития ондатры. Увеличение сетчатой зоны у самцов ондатры нами отмечено в 8-12 месяцев (табл. 15) поснатального периода, что совпадает с началом полового созревания и половым созреванием у самцов ондатры, обитающих в условиях экосистемы Южного Прибайкалья. Таблица 15 Морфометрические показатели надпочечника самцов ондатры в зависимости от возраста Показатели Толщина капсулы (мкм) Толщина (мкм) мозгового вещества (%) Толщина (мкм) коркового вещества (%) Толщина сетчатой (мкм) эоны (%) 1 25,1±0,86*** 631,5±1,60** отсутствует 27,5±1,31*** 68,0±0,50*** 298,5±5,70** * 32,0±0,50*** 76,6±1,52*** 25,3±1,31*** 140,7±1,94** * 47,2±1,31*** 81,2±3,00*** 8 80,6±1,70*** 400,4±12,10** 10 95,4±2,73 197,6±24,81** 12 97,6±4,81 224,6±22,10 27,6±0,14*** 1150,0±33,41** * 72,4±0,14*** 247,5±11,06*** 21,5±0,17*** 524,0±14,80*** 13,6±1,20*** 1244,0±31,76** * 86,4±1,20*** 141,3±10,51*** 11,3±0,61*** 656,2±18,23*** 14,2±1,02 1345,5±22,88 85,8±1,02 176,1±5,55 12,0±1,00 685,0±14,47 45,5±0,17*** 52,7±0,61*** 51,0±1,00 378,5±20,56*** 446,2±16,46*** 484,4±12,58 33,0±0,17*** 36,0±0,61*** 37,0±1,00 Примечания: процентная доля функциональных зон надпочечника дана по отношению к корковому веществу; различия с предыдущей группой достоверны: *** – при Р<0,001; ** – при Р<0,01; * – при Р<0,05 39 Толщина (мкм) клубочковой зоны (%) Толщина пучковой (мкм) зоны (%) Новорожден. 21,3±0,03** 607,3±5,63** * 71,6±0,48* 240,5±4,50** * 28,4±0,48*** 48,7±0,41*** 20,1±0,64*** 192,8±0,85** * 79,9±0,64*** отсутствует Возраст (мес.) 3 5 36,0±0,62*** 75,6±2,45*** 464,3±5,84** 468,0±44,84** * 51,2±1,00*** 37,3±1,74*** 444,3±12,85*** 780,1±37,23** * 48,8±1,00*** 62,7±1,74*** 79,3±2,72*** 163,2±8,04*** 17,8±0,53** 21,0±1,00 266,4±9,17*** 391,0±19,24** * 60,4±0,53*** 50,0±1,00*** 98,6±5,02*** 226,0±16,60** * 21,8±0,53*** 29,0±1,00*** 40 Возрастные гистохимические показатели надпочечника Гистохимически возрастные изменения выражаются в отсутствии у новорожденных самцов ондатры в структурных элементах надпочечника гликогена, который обнаруживается в 5 месячном возрасте во всех структурных элементах надпочечника. В 8 месяцев происходит снижение уровня содержания этого компонента и полное отсутствие в 10-12 месячном возрасте. Сульфатированные протеогликаны с рождения накапливаются в капсуле органа, полностью расходуясь в 10-12 месяцев. При анализе содержания белковых компонентов было отмечено учвеличение общего белка, РНК, лизина и гистидина в цитоплазме, а ДНК в ядрах гландулоцитов коры и медуллы до 5 месяцев. В этом же возрасте наблюдается появление аргинина в ядрах коркового вещества надпочечника. Количество РНК существенно увеличивается в сетчатой зоне в 10-12 месяцев, что соответствует периоду полового созревания, что по-нашему мнению связано с горонообразованием андрогенов. СЕЗОННЫЕ МОРФОФУНКЦИОНАЛЬНЫЕ ИЗМЕНЕНИЯ НАДПОЧЕЧНИКА Сезонные морфологические изменения надпочечника В конце сентября в гистоструктуре надпочечника самцов ондатры корковое вещество превалирует над мозговым (табл. 16). Таблица 16 Морфометрические показатели надпочечника самцов ондатры в зависимости от половой активности Показатели сентябрь Толщина капсулы (мкм) 94,6±4,81 Толщина (мкм) 206,6± мозгового вещества (%) 24,10*** 13,8±1,02 Толщина (мкм) 1291,4± коркового вещества (%) 22,15*** 86,2±1,02 Толщина (мкм) 158,1± клубочковой зоны (%) 5,48*** 12,2±0,96 Толщина пучковой (мкм) 667,0± зоны (%) 18,47*** 51,6±0,96 Толщина сетчатой (мкм) 466,3± эоны (%) 12,60*** 36,2±0,96 Месяц октябрь 95,4±2,73 196,6± 22,18*** 13,5±1,20 1260,2± 22,86*** 86,5±1,20*** 148,3± 12,48*** 11,8±0,61*** 656,3± 18,44*** 52,0±0,61*** 454,6± 14,46*** 36,2±0,61 ноябрь 95,4±2,73 187,8±24,81 декабрь 97,6±4,81 190,6±22,10 13,8±1,20 1174,1±31,76 14,0±1,02 1180,5±22,88 86,2±1,20*** 143,6± 10,48*** 12,2±0,61*** 606,3± 18,20*** 51,6±0,61*** 424,2± 15,46*** 36,2±0,61*** 86,0±1,02 140,1±5,56 11,7±1,00 665,0±14,47 56,3±1,00 374,4±12,58 32,0±1,00 41 Примечания: процентная доля функциональных зон надпочечника дана по отношению к корковому веществу; различия с предыдущей группой достоверны: *** – при Р<0,001; ** – при Р<0,01; * – при Р<0,05 В коре надпочечника отмечается доминирующая удельная доля пучковой зоны и наименьшая клубочковой зоны (табл. 16).Надпочечник самцов ондатры является активно функционирующим органом. На это указывает высокое содержание ядер клеток содержащих эухроматин во всех функциональных структурах органа. В конце октября-ноября существенных морфологических перестроек в гистоструктуре надпочечника не отмечено, за исключением снижения содержание функционально активных ядер клеток сетчатой зоны. В конце декабря с наступлением стадии полового покоя у самцов ондатры следует отметить некоторое увеличение пучковой и уменьшение сетчатой зоны в коре надпочечника (табл. 16). Сезонные гистохимические показатели надпочечника В начале спада половой активности в гистохимическом выражении отмечается отсутствие гликогена в цитоплазме и ядрах клеток коркового и мозгового вещества. Нейтральные глкопротеины обнаруживают себя в незначительном количестве в наружной капсуле органа. В виде умеренной реакции отмечается содержание кислых сульфатированных гликопротеинов в цитоплазме кортикоцитов. Наблюдается отсутствие сульфатированных протеогликанов и гиалурунатов. При анализе содержания белковых компонентов наблюдается незначительное содержание общего белка в наружной капсуле органа и цитоплазме гландулоцитов коркового и мозгового вещества. ДНК в виде умеренной реакции содержится в ядрах всех структурных элементов железы. Довольно интенсивная реакция отмечается на РНК в корковом веществе надпочечника. Причем уровень содержания этого компонента несколько выше в пучковой и сетчатой зоне надпочечника. Отсутствуют лизин, гистидин и аргинин. С наступлением стадии полового покоя за исключением снижения уровня содержания РНК в сетчатой и частично в пучковой зоне других изменений количественного и качественного состава углеводно-белковых соединений не отмечено. МОРФОФУНКЦИОНАЛЬНЫЙ ГИСТОГЕНЕЗ МУСКУСНОЙ ПРЕПУЦИАЛЬНОЙ ЖЕЛЕЗЫ Возрастные морфологические изменения мускусной препуциальной железы Мускусная препуциальная железа самцов ондатры состоит из двух железистых органов грушевидной формы желтого цвета расположенных в паховых областях под кожей, каждый имеющий в своем составе самостоятельный выводной проток, открывающийся в области основания Таблица 17 Морфометрические показатели мускусной препуциальной железы самцов ондатры в зависимости от возраста Показатели 1 32,8±1,35*** 39,8±3,04** 3 22,1±0,84*** 42,0±4,00*** 616,0±6,34*** ВС БС 6,5±0,15*** 6,0±0,15*** 308,8±1,01** * 138,0±0,37** * 7,0±0,10 6,4±0,10*** ВС БС ВС БС 88,2±5,04*** 72,8±4,38*** 0,8±0,01 0,8±0,02 90,3±8,08*** 79,0±4,62*** 0,8±0,01* 0,8±0,02*** Толщина капсулы (мкм) Кол-во тканевых базофилов Ср. размер Длина 100,8±3,50** ацинусов * железы (мкм) Ширина 96,2±5,30*** Средний диаметр клеток (мкм) Объем ядер клеток (мкм3) ЯЦС (ед.) 10 23,7±0,70*** 56,6±8,32*** 12 25,5±1,02 59,4±7,44 3391,0±71,63 7,3±0,03 6,8±0,10 665,6±8,72*** 1260,1±8,02*** 2360,0±18,24** * 342,0±4,70*** 626,5±5,62*** 1201,0±12,12** * 7,4±0,10*** 7,7±0,16*** 9,1±0,12*** 7,1±0,06 7,2±0,20*** 8,3±0,14*** 130,0±4,02*** 101,2±5,40*** 0,8±0,03*** 0,8±0,01 152,0±9,27*** 113,2±4,62*** 0,8±0,02*** 0,8±0,01*** 617,0±26,63 433,0±30,24 0,9±0,03 0,9±0,01 255,8±2,57*** 158,1±15,00*** 141,7±15,40*** 0,9±0,03*** 0,9±0,03*** 272,1±11,31*** 219,2±9,60*** 0,9±0,01*** 0,9±0,04** 1952,8±20,76 11,4±0,16 10,3±0,64 Примечания: ВС – верхнего слоя, БС – базального слоя; различия с предыдущей группой достоверны: *** – при Р<0,001; ** – при Р<0,01; *– при Р<0,05. 42 Новорожден. 86,7±2,65*** 56,2±9,04*** Возраст (мес.) 5 8 22,8±0,52*** 24,0±1,00*** 38,0±2,80*** 52,0±10,0*** 43 головки полового члена с левой и правой дорсо-латеральной стороны. Железы покрыты тонкой, нежной соединительнотканной капсулой. С рождения у самцов ондатры мускусная препуциальная железа имеет дефинитивное строение. По строению это простая альвеолярная железа. Паренхима органа представлена многочисленными железистыми клубочками, отделенными друг от друга тонкими прослойками соединительной ткани. Клеточный состав органа представлен двумя типами (светлыми и темными) гипрехромных клеток (табл. 17). Светлый тип клеток расположен в верхнем слое ацинуса, а темный находится на базальном слое. В дальнейшем к 1 месячному возрасту, железистые клубочки объединяются в пакеты и трансформируются к 8 месячному периоду развития в мешочки ацинарной формы (табл. 17). В 8 месяцев начинается секреторная функция железы и по своей гистологической структуре она становится похожа на сальную железу. В 10-12 месячном возрасте мускусная препуциальная железа самцов ондатры активно функционирует по голокриновому типу секреции. Возрастные гистохимические показатели мускусной препуциальной железы В секреторных клетках железы с рождения в значительном количестве выявляется гликоген. Его больше в капсуле органа и в оболочке кровеносных сосудов. Этот компонент накапливается до 3 месячного возраста, а в 5 месяцев происходит постепенное снижение уровня данного углеводного компонента до 10-12 месячного возраста. Расход гликогена выше в соединительнотканных прослойках и наружной капсуле мускусной препуциальной железы. Нейтральные гликопротеины у новорожденных щенков ондатры в незначительном количестве выявляются в наружной капсуле органа и полностью отсутствуют в клетках гландулоцитов. В 5 месячном возрасте они в большом количестве обнаруживаются в цитоплазме и ядрах гландулоцитов, полностью расходуясь к 8 месячному возрасту. Однако в соединительнотканных прослойках железы и во внутреннем слое капсулы их количество возрастает, постепенно снижаясь к 10-12 месяцам. Сульфатированные протеогликаны на раннем этапе постнатального онтогенеза обнаруживаются в соединительнотканных прослойках и наружной капсуле органа в незначительном количестве, с возрастом никак не изменяясь. В 8-месячном возрасте их количество несколько увеличивается, полностью расходуясь к 10-12 месячному возрасту. Кислые сульфатированные гликопротеины с рождения обнаруживаются в виде слабой реакции в наружной капсуле и соединительнотканных прослойках железы. В 1 месячном возрасте они постепенно накапливаются в железистых структурах до 10-12 месяцев, а в соединительнотканных прослойках и капсуле органа не выявляются с 5 месячного возраста. 44 Гиалуронаты обнаруживаются у новорожденных щенков самцов ондатры в небольшом количестве в соединительнотканных прослойках железы до 5 месячного возраста. В последующие исследуемые возрастные периоды постнатального онтогенеза они не выявляются. С рождения у самцов ондатры в секреторных клетках железы не выявляется РНК. Этот компонент появляется в 1 месячном возрасте, постепенно накапливаясь до 3 месячного возраста. Начиная с 5 месяцев, его количество уменьшается, полностью расходуясь в секреторных клетках железы в 8 месяцев. Расход РНК выше в паренхиме мускусной препуциальной железы, в соединительно-тканных прослойках и наружной капсуле органа РНК в незначительном количестве сохраняется до 10-12 месяцев. ДНК у новорожденных щенков ондатры в значительном количестве выявляется в цитоплазме и ядрах гландулоцитов секреторных отделов находящихся на периферии органа, межуточной соединительной ткани и наружной капсуле мускусной препуциальной железы. Затем в 3 месячном возрасте происходит увеличение содержания этого компонента, снижение его содержания в 5-8 месяцев. Лизин и гистидин в значительном количестве у новорожденных выявляется в наружной капсуле, умеренно окрашиваются в ядрах гландулоцитов. В 3 месячном возрасте реакция на гистидин усиливается в ядрах гландулоцитов и снижается в наружной капсуле. В 5 месячном возрасте происходит увеличение содержания обоих компонентов, затем количество лизина уменьшается в железистых структурах железы, а гистидин полностью расходуется в 8 месяцев, проявляя себя в незначительном количестве в наружной капсуле. Причем такое состояние сохраняется до 10-12 месячного возраста. Общий белок с рождения в значительном количестве содержится в соединительно-тканных прослойках и наружной капсуле железы. В цитоплазме и ядрах гландулоцитов его содержание несколько меньше. Начиная с 1 месячного возраста, содержание этого компонента в цитоплазме и ядрах гландулоцитов увеличивается, а в соединительно-тканных прослойках и наружной капсуле в 3 месяца уменьшается. С 5 до 10-12 месячного возраста происходит расходование этого компонента во всех структурных компонентах мускусной препуциальной железы. СЕЗОННЫЕ МОРФОФУНКЦИОНАЛЬНЫЕ ИЗМЕНЕНИЯ МУСКУСНОЙ ПРЕПУЦИАЛЬНОЙ ЖЕЛЕЗЫ Сезонные морфологические изменения мускусной препуциальной железы В конце сентября у самцов ондатры мускусная препуциальная железа является хорошо выраженным дольчатым образованием. Орган разделен на крупные трапециевидные доли, в которые включены многочисленные ацинусы разделенные очень тонкими соединительнотканными перегородками. Большинство выводных протоков железы заполнены сальным секретом, что свидетельствует о функциональной активности органа 45 в данный период. Функциональная активность мускусной препуциальной железы напрямую связана с функцией репродуктивной системы ондатры. В течение снижения половой активности до наступления стадии полового покоя отмечается угасание функции мускусной препуциальной железы. Об этом свидетельствует уменьшение содержания секреторных клеток с функционально активными ядрами, отсутствие в просветах выводных протоков секрета во время стадии полового покоя, уменьшение размера ацинусов (табл. 18). Таблица 18 Морфометрические показатели мускусной препуциальной железы самцов ондатры в зависимости от половой активности Показатели Сентябрь 25,6±1,00*** Период времени (мес.) Октябрь Ноябрь 26,0±1,31*** 28,4±0,80*** Декабрь 30,8±1,47 49,0±1,60*** 44,0±1,20*** 35,4±0,72*** 34,0±1,20 3246,0±33,05** * 1888,5±8,67*** 11,2±0,19*** 10,0±0,10*** 2912,3±22,42** * 1762,5±17,62** * 10,7±0,26*** 9,5±0,14*** 2429,4±43,00 ВС БС 3458,1±18,27** * 1946,6±12,40** * 11,4±0,14*** 10,1±0,16 ВС БС ВС БС 615,2±1,84*** 430,3±2,00*** 0,9±0,02*** 0,9±0,01*** 614,1±3,28*** 429,1±2,44*** 0,9±0,03*** 0,9±0,03*** 608,0±1,10*** 421,8±1,60*** 0,8±0,01*** 0,8±0,04** 572,0±3,75 391,0±4,53 0,8±0,03 0,8±0,01 Толщина капсулы (мкм) Кол-во тканевых базофилов Ср. размер Д ацинусов железы (мкм) Ш Средний диаметр клеток (мкм) Объем ядер клеток (мкм3) ЯЦС (ед.) 1410,7±11,78 10,1±0,16 9,0±0,14 Примечания: различия с предыдущей группой достоверны: *** – при Р<0,001; ** – при Р<0,01; * – при Р<0,05 Сезонные гистохимические показатели щитовидной железы В начале регрессии половой функции гликоген в небольшом количестве содержится в наружной капсуле, в междольковых и межацинарных соединительнотканных прослойках, а также в цитоплазме и ядрах гландулоцитов базального слоя. В течение спада половой активности содержание гликогена снижается, а в стадии полового покоя (конец декабря) он отсутствует в структурах мускусной препуциальной железы. В структурных элементах мускусной препуциальной железы в исследуемый период нами идентифицированы различные группы гликопротеинов: нейтральные и кислые сульфатированные. Нейтральные гликопротеины в начале снижения репродуктивной функции у самцов ондатры в незначительном количестве выявляются в наружной капсуле, 46 соединительнотканных прослойках и секреторных клетках базального слоя. Существенное количество этого компонента отмечается в гландулоцитах расположенных ближе к выводным протокам, в секрете и несколько меньше в эпителии выводных протоков. В конце ноября их содержание уменьшается в цитоплазме гландулоцитов базального слоя, а в конце декабря в гландулоцитах расположенных ближе к выводным протокам. Кислые сульфатированные гликопротеины незначительно проявляют себя в соединительнотканных структурах железы и в цитоплазме секреторных клеток. В течение исследуемого периода их содержание увеличивается в соединительнотканных прослойках и наружной капсуле и остается на одном уровне в цитоплазме гландулоцитов. Сульфатированные протеогликаны в начале спада половой активности содержатся в небольшом количестве в соединительнотканных прослойках и наружной капсуле мускусной препуциальной железы. К наступлению стадии полового покоя их количество увеличивается, что связано с изменением паренхимостромального процентного соотношения в пользу стромы, вследствие угасания функциональной активности железы. В тканях мускусной препуциальной железы самцов ондатры уровень содержания общего белка неравномерный. В начале спада половой функции он в значительном количестве содержится в наружной капсуле и соединительнотканных прослойках. Несколько меньше его в цитоплазме и ядрах секреторных клеток и эпителии выводных протоков. В паренхиме мускусной препуциальной железы самцов ондатры в начале спада половой активности РНК содержится в небольшом количестве, уровень её снижается и становится незначительным во время наступления стадии полового покоя. ДНК в начале снижения репродуктивной функции содержится в небольшом количестве в ядрах секреторных клеток базального слоя и гландулоцитах, находящихся ближе к выводным протокам. Во время стадии полового покоя уровень ДНК в ядрах едва уловим. В цитоплазме гландулоцитов в начале периода снижения половой активности в большом количестве содержатся лизин и гистидин. Аргинин в небольшом количестве содержится в ядрах гландулоцитов. Постепенно расходуясь, лизин и гистидин в незначительном количестве обнаруживаются при наступлении стадии полового покоя у самцов ондатры. Аргинин полностью расходуется в конце ноября. ВЫВОДЫ 1. Семенные канальцы самцов ондатры при рождении представлены сустентоцитами и гоноцитами. На стадии полового покоя – сустентоцитами и сперматогониями. В 5 месяцев постнатального периода гоноциты дают начало новым генерациям – сперматогониям, в 8 месяцев – первичным сперматоцитам. Намного раньше появляются просветы в канальцах семенниковой сети. Гоноциты выявляются в эпителии сети семенника у 47 щенков до 1 месячного периода развития, в последующие возрастные периоды они не обнаруживаются. Интерстициальные эндокриноциты (ИЭ) имеют два цитотипа и обнаруживаются в интерстиции гонады с рождения. Количество их с возрастом уменьшается. У половозрелых самцов ИЭ становятся однотипными. 2. Придаток семенника щенков ондатры с рождения представлен выносящими канальцами (головка), протоком придатка семенника и межуточной тканью. Эпителий выносящих канальцев и протока придатка простой низкостолбчатый. У отдельных эпителиоцитов на апикальных краях имеются реснички. С возрастом высота эпителия и диаметр протока увеличивается. Причем увеличение происходит от головки к хвосту. Особенно интенсивное увеличение вышеуказанных показателей происходит в 10-12 месяцев. Высота стереоцилий в придатке семенника ондатры уменьшается по мере приближения к хвостовой зоне придатка. Спермии идентифицируются в 10 месяцев, что свидетельствует о половом созревании в этом возрасте. В период полового покоя наблюдается существенное уменьшение указанных параметров. Спермии в состоянии полового покоя не выявляются. 3. К моменту рождения у ондатры предстательная железа, пузырьковидная железа, щитовидная железа, мускусная препуциальная железа и надпочечник имеют дефинитивное строение. Придаточные половые железы относятся к трубчато-альвеолярному типу строения и секретируют по мерокриновому типу. Мускусная препуциальная железа по всем морфологическим показателям относится к сальным железам и секретирует по голокриновому типу. Щитовидная железа принадлежит к смешаннофолликулярному типу с преобладанием мелких фолликулов. Надпочечник характеризуется с рождения отсутствием сетчатой зоны в структуре органа, которая появляется в 1 месяц постнатального периода. 4. Изменение интенсивности роста и функции семенника, придаточных половых, эндокринных желез и мускусной препуциальной железы происходит линейно: максимальный коэффициент роста отмечен в 5 месячном возрасте постнатального онтогенеза. Функциональная активность желез находится в прямой зависимости от полового созревания самцов ондатры и уровня половой активности. Наибольшая функциональная активность и динамика роста отмечена в 8-10 месяцев постнатального развития, что соответствует началу и окончанию полового созревания самцов ондатры обитающих в условиях экосистемы Южного Прибайкалья. Исключением является щитовидная железа у которой отмечается снижение функциональной активности. 5. Для внутренних половых органов самцов ондатры, эндокринных желез и мускусной препуциальной железы характерен углеводно-белковый тип секрета. Во всех железах в разные сроки постнатального онтогенеза и периодов половой активности отмечается содержание гликогена, нейтральных и кислых сульфатированных гликопротеинов, протеогликанов, 48 сиалогликопротеинов и гиалурунатов. Белковые компоненты представлены РНК, ДНК, общим и катионым белками, лизином, гистидином и аргинином. В щитовидной и мускусной препуциальной железе, надпочечнике не обнаруживается свободный катионный белок и сиалогликопротеины. 6. Мускусная препуциальная железа – видоспецифический железистый орган самцов ондатры имеющийся у данного вида животных и характеризующийся морфофункциональными особенностями. Функция мускусной препуциальной железы зависит от стадии поснатального онтогенеза и регулируется репродуктивной системой самцов ондатры. 7. Внутренние половые органы самцов ондатры, эндокинные железы и мускусная препуциальная железа имеют функциональную взаимосвязь и существуют как единый органокомплекс. 8. Уровень физиологических процессов изучаемых органов на различных стадиях постнатального развития и в зависимости от уровня половой активности является видовой особенностью данных животных. 9. Разработан способ оценки экологической обстановки в зоне экосистемы озера Байкал, где в качестве биоиндикатора предложено использовать ондатру – Патент РФ на изобретение № 2430367, зарегистрирован в Государственном реестре изобретений РФ 27.09.2011. ПРАКТИЧЕСКИЕ ПРЕДЛОЖЕНИЯ Результаты исследований могут быть использованы: – в учебных процессах при чтении лекций и проведении практических и лабораторных занятий на биологических, охотоведческих и зооинженерных факультетах высших учебных заведений; – при написании соответствующих разделов учебников и справочных руководств по эволюционной, функциональной, сравнительновидовой, половой, возрастной морфологии и эндокринологии млекопитающих и пушных, промысловых животных; – для познания закономерностей адаптациогенеза в связи с влиянием природно-климатических и половой активности в качестве нормативных критериев; – при разработки научных программ, связанных с изысканием потенциальных резервов для размножения ондатры с целью повышения популяции и увеличения охотничьих ресурсов этого вида в условиях экосистемы озера Байкал; – в эколого-физиологических исследованиях для использования ондатры в качестве биоиндикатора экологической обстановки в районе экосистемы озера Байкал. 49 СПИСОК ПУБЛИКАЦИЙ ПО ТЕМЕ ДИССЕРТАЦИИ Статьи в ведущих научных журналах, рекомендованных ВАК РФ для публикации основных результатов диссертации 1. Силкин И.И. Сезонные структурно-функциональные изменения внутренних половых органов самцов ондатры в условиях экосистемы Байкальского региона / И.И. Силкин // Вестник КрасГАУ. – 2009. – Вып. 5. – С. 103-106. 2. Силкин И.И. Зоокультура – новое направление в биотехнике размножения животных на примере ондатры экосистемы озера Байкал / А.П. Солдатов, И.И. Силкин // Труды Кубанского государственного аграрного университета. – 2009. – № 1 (2). – С. 224-225. 3. Силкин И.И. Морфо-функциональная активность щитовидной железы ондатры в постнатальном онтогенезе / Т.С. Балтухаев, И.И. Силкин // Вестник КрасГАУ. – 2009. – Вып. 10. – С. 90-93. 4. Силкин И.И. Динамика содержания углеводных компонентов в надпочечнике самцов ондатры в постнатальном периоде развития / А.П. Солдатов, И.И. Силкин // Вестник КрасГАУ. – 2009. – Вып. 11. – С. 155-158. 5. Силкин И.И. Гормональная функция щитовидной железы и гонад у самцов ондатры в период постнатального онтогенеза / И.И. Силкин, Л.Ф. Шолохов // Вестник Челябинского государственного педагогического университета. – 2009. – №. 10.2 – С. 312-319. 6. Силкин И.И. Видовые и возрастные морфо-функциональные особенности тканевых базофилов в органах половой системы самцов и самок домашних животных / Е.А. Томитова, А.П. Попов, И.И. Силкин // Вестник БГСХА. – 2010. – № 1 (18). – С. 16-22. 7. Силкин И.И. Возрастная динамика углеводных компонентов в мускусных препуциальных железах самцов ондатры / И.И. Силкин, А.П. Попов // Вестник БГСХА. – 2010. – № 1 (18). – С. 23-27. 8. Силкин И.И. Возрастная динамика некоторых белковых компонентов в мускусных препуциальных железах самцов ондатры / И.И. Силкин // Вестник КрасГАУ. – 2010. – Вып. 8. – С. 86-89. 9. Силкин И.И. Гормональная функция щитовидной железы и гонад самцов ондатры в период спада половой активности в условиях экосистемы Байкальского региона / И.И. Силкин, Л.Ф. Шолохов // Вестник КрасГАУ. – 2010. – Вып. 11. – С. 145-147. 10. Силкин И.И. Гистоморфология и содержание углеводных компонентов в мускусных препуциальных железах самцов ондатры в постнатальном онтогенезе / И.И. Силкин // Известия Смоленского государственного университета. – 2010. – № 4(12). – С. 29-37. 50 11. Силкин И.И. Возрастная динамика содержания углеводных компонентов в щитовидной железе ондатры / Т.С. Балтухаев, И.И. Силкин // Вестник КрасГАУ. – 2011. – Вып. 6. – С. 128-131. 12. Силкин И.И. Содержание углеводных соединений в щитовидной железе ондатры в зависимости от половой активности / Т.С. Балтухаев, И.И. Силкин // Вестник ИрГСХА. – 2011. – Вып. 43. – С. 100-104. 13. Силкин И.И. Функциональная активность щитовидной железы и гонад самцов ондатры в постнатальном периоде онтогенеза в условиях экосистемы Байкальского региона / И.И. Силкин // Бюллетень Восточно-Сибирского научного центра СО РАМН. – 2011. – Вып. 3(79). – ч. 2 – С. 153-156. 14.Силкин И.И. Морфологические изменения мускусных препуциальных желёз самцов ондатры и содержание в них белковых компонентов в зависимости от уровня половой активности / Т.С. Балтухаев, И.И. Силкин, Л.Ф. Шолохов // Известия Иркутского государственного университета. – 2011. – Т. 1. - №4. – С. 129-135. 15.Силкин И.И. Гистологические изменения и содержание белковых компонентов в мускусных препуциальных железах самцов ондатры на этапах постнатального онтогенеза / И.И. Силкин, А.П. Попов // Научный журнал КубГАУ. – 2012. – № 78(04). – C. 1–12. http://www/ej.kubagro.ru/2012/04/pdf/18.pdf. 16. Силкин И.И. Морфофункциональная характеристика мускусной препуциальной железы самцов ондатры в зависимости от половой активности / И.И. Силкин // Вестник Оренбургского государственного университета. – 2012. – № 10 (146). – С. 129-133. Монография 17. Силкин И.И. Гистофизиология внутренних половых органов самцов ондатры / И.И. Силкин, А.П. Попов. – Иркутск: Изд-во Иркутского государственного университета, 2009. – 127 с. Патент 18. Силкин И.И. Способ оценки экологической обстановки в зоне экосистемы озера Байкал / И.И. Силкин, А.П. Попов // Патент РФ на изобретение № 2430367, зарегистрирован в Государственном реестре изобретений РФ 27.09.2011. Статьи, опубликованные в сборниках научных трудов и материалах международных и всероссийских конференций, других периодических изданиях 19. Силкин И.И. Возрастные изменения морфометрических показателей семенников ондатры в постнатальном онтогенезе / И.И. Силкин // 51 Теоретические и практические аспекты вет. медицины: Мат. межрегион. науч. конф. – Улан-Удэ, 2001. – С. 88-89. 20. Силкин И.И. Макро- микроморфология и гистохимические показатели пузырьковидной железы ондатры / И.И. Силкин // Объединенный научный журнал. – 2002. – №26/49 – С. 45-46. 21. Силкин И.И. Морфо-функциональная характеристика семенников ондатры дельты р. Селенга / И.И. Силкин, А.П. Попов // Структура и функционирование экосистем Байкальской Сибири: Мат. регион. научно-практич. конф. – Улан-Удэ, 2003. – С. 91-92. 22. Силкин И.И. Гистоморфология и содержание некоторых углеводных компонентов в предстательной железе ондатры / И.И. Силкин, А.П. Попов // Актуальные вопросы вет. медицины мелких домашних животных: Мат. 5 Всероссийской конф. – Екатеринбург, 2003. – С. 123-125. 23. Силкин И.И. Динамика возрастных изменений некоторых углеводных компонентов в половых органов самцов ондатры / И.И. Силкин // Мат. научно-практич. конф., посв. 70-летию ИрГСХА. – Иркутск, 2004.– С. 120-124. 24. Силкин И.И. Морфо-функциональный гистогенез гонад новорожденных щенков ондатры / И.И. Силкин // Актуальные аспекты экологической, сравнительно-видовой, возрастной и эксперимент. морфологии: Мат. международ. научно-практич. конф., посв. 100летию проф. Суетина В.Я. – Улан-Удэ, 2004. – С. 173-174. 25. Силкин И.И. Сезонные морфо-функциональные изменения придатка семенника / И.И. Силкин // Актуальные проблемы АПК: Мат. регион. научно-практич. конф. – Иркутск, 2005. – С. 49-50. 26. Силкин И.И. Структурно-функциональный гистогенез пузырьковидной железы ондатры / И.И. Силкин // Проблемы устойчивого развития АПК: Мат. научно-практич. конф. – Иркутск, 2006. – С. 60-65. 27. Силкин И.И. Структурно-функциональные изменения в семенниках ондатры в зависимости от сезона года / И.И. Силкин // Вестник Тюменской гос. сельхоз. академии. – 2007. – №1/2. – С. 164-165. 28. Силкин И.И. Структурно-функциональные особенности семенников ондатры экосистемы озера Байкал / И.И. Силкин // Роль молодых ученых в развитии науки: Мат. 2 научно-практич. конф. – Великие Луки, 2007. – С. 23-26. 29. Силкин И.И. Морфогистохимические изменения щитовидной железы при тиоцианатном гипотериозе / Т.А. Балтухаева, Н.К. Шотников, Т.С. Балтухаев, И.И. Силкин // Актуальные вопросы развития регионального АПК: Мат. научно-практич. конф. – Иркутск, 2007. – С. 8-9. 52 30. Силкин И.И. Топография и морфометрические показатели щитовидной железы ондатры в постнатальном онтогенезе / Т.С. Балтухаев, И.И. Силкин // Совместная деятельность сельхоз. производителей и науч. организаций в развитии АПК Центральной Азии: Мат. международной научно-практич. конф. – Иркутск, 2008. – С. 12-18. 31. Силкин И.И. Топография и морфометрические показатели мускусной железы ондатры в постнатальном онтогенезе / И.И. Силкин // Совместная деятельность сельхоз. производителей и науч. организаций в развитии АПК Центральной Азии: Мат. международной научно-практич. конф. – Иркутск, 2008. – С. 244-249. 32. Силкин И.И. Топография и морфометрические показатели надпочечников ондатры в постнатальном онтогенезе / А.П. Солдатов, И.И. Силкин // Совместная деятельность сельхоз. производителей и науч. организаций в развитии АПК Центральной Азии: Мат. международной научно-практич. конф. – Иркутск, 2008. – С. 249-255. 33. Силкин И.И. Структурная организация мускусной железы ондатры / И.И. Силкин // Актуальные вопросы инвазионной и инфекционной патологии животных: Мат. международной конф. – Улан-Удэ, 2008. – С. 97-98. 34. Силкин И.И. Анатомо-гистологическая характеристика щитовидной железы ондатры / Т.С. Балтухаев, И.И. Силкин // Актуальные вопросы инвазионной и инфекционной патологии животных: Мат. международной конф. – Улан-Удэ, 2008. – С. 102-104. 35. Силкин И.И. Макро-, микроморфология надпочечников ондатры / А.П. Солдатов, И.И. Силкин // Актуальные вопросы инвазионной и инфекционной патологии животных: Мат. международной конф. – Улан-Удэ, 2008. – С.104-106. 36. Силкин И.И. Структурная организация щитовидной железы новорожденных щенков ондатры / Т.С. Балтухаев, И.И. Силкин // Состояние и перспективы обеспечения ветеринарного благополучия Восточной Сибири: Мат. международной научно-практич. конф. – Чита, 2008. – С. 69-73. 37. Силкин И.И. Анатомо-гистологическая характеристика мускусной железы ондатры в ранний постнатальный период онтогенеза / И.И. Силкин // Состояние и перспективы обеспечения ветеринарного благополучия Восточной Сибири: Мат. международной научнопрактич. конф. – Чита, 2008. – С. 193-197. 38. Силкин И.И. Изопропиловый спирт для проводки – оптимальный метод в гистотехнике / И.И. Силкин, О.П. Ильина // Состояние и перспективы обеспечения ветеринарного благополучия Восточной Сибири: Мат. международной научно-практич. конф. – Чита, 2008. – С. 197-202. 53 39. Силкин И.И. Структурная организация надпочечника ондатры в период раннего постнатального онтогенеза / А.П. Солдатов, И.И. Силкин // Состояние и перспективы обеспечения ветеринарного благополучия Восточной Сибири: Мат. международной научнопрактич. конф. – Чита, 2008. – С. 223-227. 40. Силкин И.И. Гистологическое строение и углеводные компоненты щитовидной железы новорожденных щенков ондатры / Т.С. Балтухаев, И.И. Силкин // Актуальные вопросы электрофизиологии и незаразной патологии животных: Мат. международной научно-практич. конф. посв. 70-летию зав. каф. терапии и клинической диагностики Ю.А. Тарнуева. – Улан-Удэ, 2009. – Ч. 2 – С. 8-11. 41. Силкин И.И. Ондатра как представитель зоокультуры экосистемы Байкальского региона / И.И. Силкин // Актуальные вопросы электрофизиологии и незаразной патологии животных: Мат. международной научно-практич. конф. посв. 70-летию зав. каф. терапии и клинической диагностики Ю.А. Тарнуева. – Улан-Удэ, 2009. – Ч. 2 – С. 81-83. 42. Силкин И.И. Морфология надпочечников новорожденных щенков ондатры и содержание в них углеводных компонентов / А.П. Солдатов, И.И. Силкин // Актуальные вопросы электрофизиологии и незаразной патологии животных: Мат. международной научнопрактич. конф. посв. 70-летию зав. каф. терапии и клинической диагностики Ю.А. Тарнуева. – Улан-Удэ, 2009. – Ч. 2 – С. 86-89. 43. Силкин И.И. Возрастной структурно-функциональный гистогенез внутренних половых органов самцов ондатры экосистемы 53 Байкальского региона / И.И. Силкин // Проблемы зооинженерии и ветеринарной медицины: Сб. научных трудов. – Харьков, 2009. – Вып. 19. – Ч. 2. – Том 2. – С. 70-79. 44. Силкин И.И. Модификация метода тетразониевого азосочетания (по Даниелли) с использованием прочного синего Б (по Берстону) / И.И. Силкин, Б.Я. Власов // Вестник Полтавской государственной аграрной академии. – Полтава, 2009. – № 2 (53). – С. 68-69. 45. Силкин И.И. Строение и некоторые гистохимические показатели мускусной железы ондатры в ранний постнатальный период онтогенеза / И.И. Силкин // Актуальные проблемы ветеринарной морфологии: Мат. международной науч. конф. посв. 90-летию каф. анатомии животных СПбГАВМ. – С-Пб., 2009. – С. 77-81. 46. Силкин И.И. Использование ондатры в качестве нового тест-объекта для оценки экологической обстановки районов Прибайкалья / А.П. Солдатов, И.И. Силкин // Охрана и рациональное использование животных и растительных ресурсов: Мат. международной научно- 54 практич. конф. посв. 60-летию фак. охотоведения им. В.Н. Скалона. – Иркутск, 2010. – С. 525-529. 47. Силкин И.И. Анатомо-топографические особенности половой системы самцов ондатры / И.И. Силкин, А.П. Попов // Морфология XXI столетия: Мат. международной научной конф. посв. 100-летию со дня рождения проф. Г.О. Гиммельрейха и 90-летию каф. анатомии им. академика В.Г. Касьяненко. НУБП Украины. – Киев, 2010. – С. 206214. ИСПОЛЬЗОВАННЫЕ СОКРАЩЕНИЯ И АБРРЕВИАТУРЫ ИЭ – интерстициальные эндокриноциты; ЛГ – лютеинизирующий гормон (лютеотропин, лютропин); ФСГ – фолликулостимулирующий гормон (фоллитропин); Т3 – трийодтиронин; Т4 – тироксин; Т3св – трийодтиронин связанный; Т4св – тироксин связанный;