полученных по различным методикам. При этом БИОЛОГИЧЕСКИЕ НАУКИ
реклама

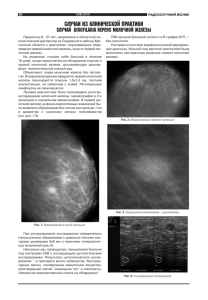
БИОЛОГИЧЕСКИЕ НАУКИ Литература полученных по различным методикам. При этом оказалось, что нативный антиген гриба Tr. faviforme обладает специфической активностью в РА и ИФА, слабой преципитирующей активностью, выявленной в РП по Оухтерлони. Полные анти гены гриба Tr. faviforme обладают высокой спе цифической активностью в РА и ИФА. Пре ципитирующие свойства у полных антигенов не выявлены. Наиболее активными антигенными свойствами в ИФА при выявлении противотри хофитийных антител обладает полный антиген по методу БуавенуМесробеану. 1. Равилов, Р.Х. Проблемы борьбы с инфекциями мелких до машних животных // Актуальные вопросы ветеринарной медицины: мат. сибирской межд. научн.практ. конф. Но восибирск, 2004. С. 38–40. 2 . Овчинников, P.C. И зучение изменчивости морфологи ческих характеристик дерматофитов / Р.С. Овчинников, К.П. Летягин, А.Н. Панин // Доклады РАСХН. 1999. № 5. С. 37–39. 3 . Кухар, Е.В. Использование иммуноферментной тестсис темы на основе поликлональных антител в диагностике трихофитии крупного рогатого скота / Е.В. Кухар, К.К. Му канов // Естествознание и гуманизм / Сибирский государ ственный медицинский университет. Томск: ООО «Вайар». 2005. № 2. С. 99–101. 4 . Методы экспериментальной микологии: справочник. К.: Наукова думка, 1982. С. 122–124, 240–249, 390–415. Фенотипы тучных клеток молочной железы и яичника беременных крольчих ­Г. Калякина, аспирантка, Оренбургский ГАУ Успешность работы по воспроизводству жи вотных всецело зависит от знания структуры и понимания процессов, протекающих в органах репродуктивной системы самок. Структурно функционально органы репродукции лабиль ны, реагируют на изменения гомеостаза и воз действие факторов внешней среды реорганиза цией на молекулярном, клеточном и тканевом уровнях. Регуляция деятельности яичника и молочной железы осуществляется на нейроэндокринном (ядра гипоталамуса, аденогипофиз, эпифиз, яич ник, матка) и местном дистантном уровнях [1]. В морфофункциональных микрорайонах ор ганов регуляцию функций в ответ на изменение гомеостаза обеспечивают тучные клетки (ТК) посредством выработки биологически активных веществ (БАВ) [2, 3, 4]. Цель настоящего исследования: изучить ха рактеристики популяций ТК в молочной железе и яичнике крольчих в период беременности и выявить закономерности в изменении их феноти пов в зависимости от функциональной активно сти структур микрорайонов. Материал и методы. Объектом исследования служили молочные железы и яичники 17 бере менных крольчих. Для гистологического иссле дования забирали пробы объемом 0,5 см3. Часть гистопроб фиксировали в жидком азоте. Изготов ляли криостатные срезы толщиной 10–12 мкм и окрашивали по стандартной методике толуидино вым синим. Количественную информацию о тучноклеточ ных популяциях получали в ходе морфометричес ких исследований при использовании винтового окулярмикрометра МОВ–1–15 1350. В отдель ном образце ткани измерения каждого показате ля осуществляли не менее чем в 15 полях зрения каждого объекта. Индекс фенотипа популяции (ИФП) тучных клеток определяли по формуле: ИФП = Д/Н, где Д – количество дегранулирующих тучных клеток; Н – количество недегранулирующих тучных клеток. Результаты и обсуждение. Выявлено, что попу ляции тучных клеток в молочной железе и яични Рис. 1 – Фенотипы ТК в молочной железе и яичнике беременных крольчих: А – фенотип I (молочная железа); Б – фенотип II (яичник); В – фенотип III (яичник) 104 БИОЛОГИЧЕСКИЕ НАУКИ Итак, тучные клетки первого фенотипа лока лизовались в тканевых микрорайонах, отвечаю щих за трофику и являющихся материалом для построения функционально зрелых структур органа. В области формирующихся и сформирован ных железистых долек молочной железы, по пе риферии желтого тела и растущих фолликулов яичника отмечали локализацию тучных клеток третьего типа, которые уже вступили в фазу де грануляции. Для этих клеток характерны ядер ный белковый синтез (подготовка к следующе му секреторному циклу) и выведение гранул. Ак тивное действующее вещество вышедших в межклеточное пространство гранул воздейству ет на состояние гистогематического барьера сте нок сосудов гемомикроциркуляторного русла, регулирует цикличность секреторного цикла в молочной железе, рост фолликулов и синтез гор монов – в яичнике. В междольковых соедини тельнотканных трабекулах и протоках молочной железы, а также в зоне примордиальных фолли кулов яичника данного типа клеток не обнару жили. Тучные клетки второго фенотипа наблюдали во всех тканевых микрорайонах органов и харак теризовали как «переходный» между клетками первого и третьего типов. Таким образом, тучные клетки образуют в молочной железе и яичнике крольчих гетероген ные и пластичные популяции, что согласуется с данными Н.А. Юриной, А.И. Радостиной (1990), Schwartz L. (1993). Их численность и состав как в целом, так и в отдельных участках органов регулируются на различных уровнях. Фенотипи ке гетерогенны по фенотипу. В каждом органе в один и тот же функциональный период выявля ется несколько типов клеток. 1. Клетки крупные, ядра темные, с высоким содержанием конденсированного хроматина, цитоплазма темная, не содержит гранул (рис. 1а). 2. Клетки средних размеров, ядра светлые, насыщенные деконденсированным хроматином, цитоплазма светлая, содержит гранулы, выход ко торых незначителен или еще не начался (рис. 1б). 3. Клетки мелкие, ядра содержат глыбки кон денсированного хроматина, цитоплазма светлая, гранулы лежат в цитоплазме и рядом с клеткой (рис. 1в). Во всех фенотипах диаметр ТК в молочной железе был больше диаметра ТК соответствую щего фенотипа в яичнике. Разница в размерах составила 1,5–1,6 раза (табл. 1). Во всех ткане вых микрорайонах исследуемых органов популя ция тучных клеток была неоднородна и представ лена двумятремя типами. Процентное соотношение фенотипов показа но в таблице 2. Клетки первого типа характеризовались высо кой синтетической активностью, в их цитоплаз ме регистрируется активизация белоксинтезиру ющих систем, происходит накопление белков и БАВ. Этот тип клеток выявляли в молочной же лезе в междольковых соединительнотканных тра бекулах, вокруг междольковых протоков, в жиро вых дольках, а в яичнике – под белочной оболоч кой в зоне примордиальных фолликулов. Рядом с растущими фолликулами и желтым телом яич ника, а также вблизи альвеол молочной железы клетки данного типа не обнаруживали. 1. Морфометрические показатели тучных клеток разных фенотипов в молочной железе и яичнике беременных крольчих (мкм) Фенотип ТК Показатель Диаметр клеток Степень дегрануляции I молочная железа 8,682±0,023 II яичник молочная железа 5,824±0,017 6,763±0,019 нет III яичник молочная железа яичник 4,187±0,041 4,763±0,019 3,097±0,034 незначительная значительная 2. Соотношение фенотипов ТК молочной железы и яичника беременных крольчих Показатель Микрорайон ИФП Фенотип ТК, % I II III 0 0,23 1,71 3,76 58 53 11 0 42 28 26 21 0 19 63 79 0,625 1,0 1,0 41 0 0 23 46 48 36 54 52 Молочная железа Междольковые соединительнотканные трабекулы и протоки Жировые дольки Формирующиеся железистые дольки Альвеолы сформированных железистых долек Яичник Примордиальные фолликулы Растущие фолликулы Желтое тело беременности 105 БИОЛОГИЧЕСКИЕ НАУКИ Литература ческие признаки тучных клеток во многом зави сят от фазы синтеза биологически активных ве ществ и от факторов микроокружения. Выявлен ная разница диаметров ТК фенотипов молочной железы и яичника (в 1,5–1,6 раза) в одинаковые функциональные периоды обусловлена, на наш взгляд, разными источниками происхождения ТК и структур исследуемых органов. Фенотипическая пластичность ТК, очевидно, обеспечивает оптимальное осуществление локаль ными популяциями ТК специфических функ ций. 1. Metcalfe, D.D., Baram, D., and Mekori Y. Mast cells, Physiol Rev., 1997. V. 77. P. 1033–1079. 2 . Языков, В.Л. Секреторные механизмы и секреторные продук ты тучных клеток. Морфология, 1999. Т. 115. Вып. 2. С. 64–72. 3 . Арташян, О.С. Роль системного подхода в изучении системы тучных клеток в живых организмах // Философия и наука: мат. 2й межвуз. науч.практ. конф. аспирантов и соискате лей «Философия и наука». Екатеринбург. 2003. С. 3–5. 4 . Craig, S.S., Irani, A.M., Metcalfe, D.D. and Schwartz, L.B. Ultrastructural localization of heparin to human mast cells of the MCTC and MCT types by labelling with antithrombin Illgold. Lab. Invest. 1993. V. 69. P. 552–561. 5 . Ю рина, Н.А. М орфофункциональная гетерогенность и взаимодействие клеток соединительной ткани / Н.А. Юри на, А.И. Радостина. М.: Издво УДН, 1990. 6 . Schwartz L. Heterogeneity of human mast cells. In: The Mast Cell in health and Disease. New York. Dekker. 1993. P. 219–236. Изучение бурого медведя в природных условиях Оренбургской области А. Латыпов, аспирант, Оренбургский ГАУ Экологофаунистические исследования буро го медведя (Ursus arctos L., 1758) в природных условиях Оренбургской области являются абсо лютно новыми, хотя данный вид не является новым для Южного Урала. Бурый медведь как объект исследований все гда привлекал внимание множества ученых и натуралистов. Первыми исследователями терио фауны Оренбургской области являются П.С. Пал лас, Э.А. Эверсманн, П.И. Рычков. В книгах этих авторов [1, 2, 3] приводится описание соста ва животного мира Южного Урала, в частности, встречаются упоминания о встречах бурого мед ведя в лесных массивах Уральских гор. Я.Н. Даркшевич, проводя систематическую классификацию териофауны Чкаловской (Орен бургской) области, отмечает заходы бурого мед ведя на территорию Троицкого (Тюльганского) района из соседней Башкирии, таким образом, ставя под вопрос постоянное обитание медведя в центральных районах северной части области [4]. Говоря об общем и частном изучении медведя в пределах нашей страны, невозможно не упомя нуть о деятельности В.С. Пажетнова. Он прово дит разделение бурого медведя евроазиатской зоны России на семь подвидов. Согласно его классификации, на территории Оренбургской области и сопредельных регионов обитает сред нерусский бурый медведь. Результатом иссле дований Пажетнова стали многочисленные публикации, в которых он дает частное и общее описание биологии, экологии и этологии бурых медведей [5], особое внимание при этом уделяет вопросам методологии [6]. Бурый медведь является особо ценным в хозяй ственном отношении видом, в связи с чем подле жит ежегодному государственному учету. В отрас ли охотничьего хозяйства Российской Федерации, в т.ч. в Оренбургской области, учет численности бурого медведя проводится по утвержденной ме тодике [7]. Однако некоторые исследователи счи тают данную методику несовершенной [6]. Н.К. Верещагин [8] описывает особенности морфологии, географической изменчивости, гео логической истории бурого медведя, а также воп росы охраны и хозяйственного использования его ресурсов. В.П. Теплов в ходе исследований в северном Приуралье выявил прямую связь увеличения гибели домашнего скота с урожаем ягод. «Так, в годы обильного урожая от медведей погибало в среднем 0,4 головы домашнего скота, при сред нем – 4, а в годы плохого урожая – 15. Таким образом, при отсутствии ягод медведи нападали на скот в 38 раз чаще, чем обычно» [9]. Большой вклад в изучение териофауны Орен бургской области внесла В.Н. Руди. Она приво дит сводные данные о распространении бурого медведя на территории Оренбургской области [10]: «В XVIII–XIX вв. медведи обитали по р. Бу зулук (Паллас, 1788; Зарудный, 1897; Эверсманн, 1850). С середины XIX в. и до конца XX столетия медведи в области не встречались. В последние годы стали появляться в Кувандыкском, Сарак ташском и Тюльганском районах. Современная южная граница его распространения проходит по южноуральской дубравной лесостепи. Следы медведя были нами обнаружены во время экс педиции в 1970 г. в отрогах Южного Урала по хребту ДзяуТюбе. Неоднократно отмечается по явление медведей в окрестностях села Каипово Кувандыкского района». Опираясь на ежегодные учетные данные ГУ «Управление объектами животного мира и 106