Формирование ориентационной настройка простых клеток
реклама

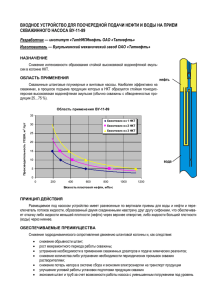
А.М. Левинов Формирование ориентационной настройка простых клеток зрительной коры Ключевые слова: простые клетки, ориентация, обучение, механизм обучения Краткое содержание. В работе показывается, что простые клетки зрительной коры могут формироваться в результате обучения. Использование идеи обучения снимает ряд противоречий, присущих представлениям о формировании этих клеток, и объясняет некоторые явления, связанные с их функционированием. С обучением увязывается также процесс возникновения в зрительной коре областей расположенных рядом одинаково ориентированных простых клеток. Проблемы, связанные с простыми клетками зрительной коры. Много лет назад Hubel и Wiesel (1962, 1968) обнаружили, что некоторые нейроны зрительной коры избирательно реагируют на определенную ориентацию прямолинейного края объекта или прямолинейной полоски. С тех пор исследование этих нейронов (простых клеток) и связанных с ними механизмов стало одной из наиболее интенсивно разрабатываемых областей нейрофизиологии. A B S C Рис. 1. Расположение ON и OFF нейронов НКТ, соответствующее рецептивному полю простой клетки зрительной коры. Незаполненные круги – ON нейроны, заштрихованные OFF нейроны. Область B – совокупность ON нейронов, посылающих сигналы на одну и ту же простую клетку; A и C зоны OFF нейронов, прилегающих к области B; S – простая клетка. Рассматривая связи между простыми клетками и нейронами наружного коленчатого тела (НКТ), Hubel и Wiesel выяснили, что ON нейроны НКТ, посылающие сигнал на одну и ту же простую клетку, функционально организованы так, как будто они расположены вдоль отрезка прямой. К ним примыкают OFF клетки НКТ, формируя зоны торможения (рис. 1). Такая картина могла бы возникнуть, если бы ON и OFF нейроны были расположены в НКТ примерно так, как показано на рисунке 2. 2 Рис. 2. Заштрихованные круги OFF клетки НКТ, незаштрихованные ON клетки НКТ; пояснения в тексте. Но оказалось, что ON и OFF клетки расположены в НКТ так, как показано на рисунке 3: каждый ON нейрон окружен OFF нейронами, образующими OFF зону. Рис. 3. Нейроны НКТ: заштрихованные кольца OFF зоны ON нейронов, круги внутри заштрихованных колец ON нейроны. Чтобы объяснить найденное несоответствие, Hubel и Wiesel (1962, 1968) предположили, что клетки НКТ и простые клетки связаны так, как показано на рисунке 4. Hubel и Wiesel полагали, что такая структура расположения клеток объясняет взаимосвязь, показанную на рисунке 1. k l m n E A B C D Рис. 4. A, B, C, D нейроны НКТ вместе с их тормозящим окружением: заштрихованные кольца OFF окружение; внутренние круги ON клетки; E простая клетка зрительной коры, на которую поступают сигналы от соответствующих ON нейронов НКТ; k, l, m, n промежуточные нейроны. 3 Эта схема взаимосвязи не всем представлялась удовлетворительной. Были предложены модели, в которых простые клетки возникали в результате развития связей и изменения синаптической проводимости нейронов зрительной коры (von der Malsburg, 1973; Tanaka, 1991; Swindale, 1996; Adorján et al., 1999; Tavazoie, Reid, 2000; Ernst et al., 2001; Teich, Qian, 2003; Ringach, 2004). Получающиеся в таких моделях структуры были в определенной мере близки к наблюдаемым. Но используемые в них предположения об исходных взаимосвязях нейронов зрительной коры в существенной мере отличались от полученного в экспериментах. Либо требовали большого участия врожденных характеристик. В то же время в экспериментах было показано, что чувствительность к ориентации зрительного стимула присуща части нейронов НКТ (Smith et al., 1990; Zhou et al., 1995; Xu et al., 2002). Исходя из этого, были предложены модели, в которых предполагалось, что какая-то чувствительность к ориентации есть уже в НКТ, а в зрительной коре она лишь усиливается (Somers et al., 1995; Ben-Yishai et al., 1995; Vidyasagar et al., 1996). С другой стороны, представление о связи клеток НКТ и V1, которое предложили Hubel и Wiesel (1962, 1968), было в какой-то мере подтверждено экспериментально. Было статистически показано, что ON клетки НКТ посылают сигналы на простые клетки, а их OFF окружение на OFF окружение простых клеток (Tanaka, 1983; Reid, Alonso, 1995; Alonso et al., 2001). Эти результаты привели к тому, что многие исследователи стали принимать схему связи клеток НКТ и V1, которую предложили Hubel и Wiesel. Таким образом, в одних моделях авторы приписывают настройку ориентации только простым клеткам области V1 зрительной коры, в других еще и нейронам из НКТ (Miller, 1994; Troyer et al., 1998; McLaughlin et al., 2000; Wielaard et al., 2001; Troyer et al., 2002; Miller, 2003; Lund et al., 2003; Shapley et al., 2003; Tao et al., 2004; Ursino, La Cara, 2004). Существующие модели объясняют многие факты, связанных с функционированием простых клеток, но некоторые авторы отмечают, что окончательные выводы пока делать рано, и что механизмы, лежащие в основе явления, пока не очень понятны (Swindale, 1996; Carandini, Ringach, 1997; Chung, Ferster, 1998; Wong, 1999; Ferster, Miller, 2000; Casagrande et al., 2002; Troyer et al., 2002; Shapley et al., 2003; Carriera-Perpiñán, Goodhill, 2004; Ursino, La Cara, 2004; Ringach, 2004). Поскольку многие исследователи в своих представлениях исходят из схемы, которую предложили Hubel и Wiesel, то нужно рассмотреть некоторые из присущих ей противоречий. Одно из них связано с самим существованием продолговатых ON рецептивных полей у простых клеток (рис. 1). а b Рис. 5. a. Множество ON и OFF нейронов НКТ, которое должно оптимально реагировать на горизонтальную полоску; b. То же множество с наложенной горизонтальной полоской. 4 Чтобы показать проблему, вернемся к предполагаемому взаимному расположению ON и OFF клеток в НКТ (рис. 3). Выберем множество нейронов, оптимально реагирующих на предъявление горизонтальной полоски (рис. 5.а). Легко видеть, что при предъявлении любой такой полоски, даже оптимально возбуждающей ON клетки (рис. 5.b), в этой структуре будут зоны торможения. Это противоречит экспериментальным данным о структуре зон возбуждения и торможения простой клетки (рис. 1). Еще одно противоречие. Схема, показанная на рисунке 4, предполагает переход от НКТ к коре с помощью промежуточных синапсов. Экспериментальные данные говорят о моносинаптичности связи нейронов НКТ с простыми клетками (Tanaka, 1983; Reid, Alonso, 1995; Usrey et al., 2000; Alonso et al., 2001). Чтобы показать еще одну проблему, связанную со схемой, которую предложили Hubel и Wiesel, вернемся к рисунку 3 и рассмотрим предлагаемый переход из НКТ на простые клетки. Схема Hubel'а и Wiesel'а предполагает, что в НКТ нейроны группируются вдоль прямолинейных полосок определенной длины. Затем каждая такая группа отображается на простую клетку области V1 (рисунок 6: чтобы не загромождать чертеж, полосы на этом рисунке содержат всего по три ON нейрона с окружающей их OFF областью). S T U Рис. 6. Схема отображения нейронов НКТ на простые клетки, следующая из идеи Hubel'а и Wiesel'а. Заштрихованные кольца OFF нейроны НКТ, внутренние незаштрихованные круги ON нейроны; прямоугольники, наложенные на нейроны НКТ − прямолинейные полоски; S, T, U простые клетки зрительной коры, на которые поступают сигналы от соответствующих ON нейронов НКТ. Проблема в самой возможности реализовать такую идею. Чтобы собрать ON клетки, лежащие в НКТ на одном и том же отрезке прямой, нужно сначала выяснить, какие из них лежат на одной прямой. Поэтому в мозгу должны быть механизмы, которые устанавливают, какие ON нейроны НКТ лежат на одной и той же прямолинейной полоске данной длины и ориентации. Должны быть механизмы, которые группируют нейроны по полоскам и собирают сигналы идущий от каждой группы. 5 Существование таких механизмов маловероятно. Но сама эта идея встречается достаточно часто. Например, Adorján et al., (1999) предполагают, что простым клеткам известно, лежат ли они на одной прямой; Shouval et al., (2000) считают, что простые клетки знают, какие из них одинаково ориентированны и лежат на одной оси. Идею знания клетками своего взаимного расположения использовали Hubel и Wiesel для объяснения структуры сложных клеток коры. Отображение из НКТ на простые клетки зрительной коры, порождающее ориентационное предпочтение, пока нельзя считать объясненным: не была доказана возможность отображения из НКТ на простые клетки, соответствующего рисунку 4; существование схемы, которую предложили Hubel и Wiesel, была показана не прямым экспериментом, а только как результат статистической обработки. Поэтому желательно рассмотреть возможность таких схем формирования простых клеток, которые отличны от предложенного Hubel'ем и Wiesel'ем. Ниже рассматривается одна из возможностей решения проблемы и описывается механизм, учитывающий не только генетически заложенные свойства нервной системы, но и те, которые получаются в результате обучения. Конечно, нет уверенности, что в формировании простых клеток зрительной коры работает именно такой механизм. Но очевидно, что при создавшейся ситуации нужны альтернативы существующей интерпретации экспериментальных данных. Формирование простых клеток зрительной коры в процессе обучения. При переходе от биполярных клеток сетчатки к ганглиозным и дальше к НКТ на каждой клетке последующего слоя сходятся окончания множества клеток предыдущего, и это множество по форме близко к кругу. При переходе от НКТ к простым клеткам зрительной коры закономерность нарушается: считается, что на каждую простую клетку приходят сигналы от области НКТ, форма которой показана на рисунке 1. Это отклонение представляется странным, и ниже предлагается схема формирования простых клеток, в которой такого нарушения нет. A S Рис. 7. Круг A − область сетчатки, которая содержит все ганглиозные ON клетки сетчатки, посылающие сигнал на простую клетку из V1; круги внутри области A − ганглиозные клетки, S − простая клетка в V1. Как показали Chapman и Gödecke (2000), в формировании простых клеток зрительной коры основная роль принадлежит ON клеткам НКТ, а роль OFF клеток пренебрежимо мала. Исходя из этого, в предлагаемой схеме обучения используются только возбуждающие нейроны: предполагается, что на простые клетки сигнал поступает только от ON клеток. Существование торможения учитывается, но об этом будет сказано позже. 6 Вообще говоря, неясно, насколько важно, что ON нейроны, несущие возбуждение на простые клетки, выходят из НКТ. В описываемом механизме предполагается, что НКТ играет только вспомогательную роль. Поэтому дальше для простоты изложения рассматривается отображение ON ганглиозных клеток сетчатки непосредственно на простые клетки зрительной коры. При этом предполагается, что (рис. 7) на простые клетки зрительной коры приходят сигналы только от ганглиозных ON нейронов сетчатки; совокупность ON нейронов сетчатки, посылающих сигналы на простую клетку зрительной коры, образует (приблизительно) круг. Дальше описывается механизм, в силу которого через какое-то время обучения сигналы на простую клетку будут проходить только от нейронов определенной полосы внутри этого круга (рис. 8). Идея опирается на предположение, что изменение синаптической проводимости некоторых нейронов происходит в результате обучения. Это внешне похоже на выдвинутую Hebb'ом (1949) идею синаптических изменений, возникающих при функционировании нервной системы. Но, как будет видно дальше, существенно от нее отличается. Ниже формулируются предположения, которые используют механизм обучения и позволяют сформировать у простых клеток V1 оптимальную реакцию на полоски определенной ориентации. A R S Рис. 8. Круг A − область сетчатки, внутри которой находятся все ганглиозные ON клетки, посылающие сигнал на простую клетку, S − простая клетка области V1, круги внутри A − ганглиозные ON клетки, прямоугольник R − полоска, на которую должна обучиться реагировать простая клетка. Чтобы показать возможность формирования отображения, показанного на рисунке 8, дальше вводится обучение синапсов, через которые на простые клетки поступают сигналы от ON нейронов. Предполагается что обучение происходит на ранних этапах развития организма. Когда глаз уже может двигаться, но еще очень медленно и без крупноамплитудных скачков. Для синапса ON клетки сетчатки с простой клеткой слоя V1 зрительной коры предполагается, что если через такой синапс в какой-то момент поступает возбуждающий сигнал, то синапс переходит в состояние ожидания; состояние ожидания длится столько, сколько нужно для фиксации глаза на объекте, его перевода в новое положение и некоторого времени рассматривания объекта в новом положении; 7 если в течение промежутка ожидания на этот синапс снова поступает возбуждение, то он увеличивается и в дальнейшем более интенсивно передает поступающий на него возбуждающий сигнал; если в течение промежутка ожидания сигнал не поступает, то с синапсом, бывшим в состоянии ожидания, ничего не происходит (то есть если мышца случайно переместит глаз не в ту сторону, куда нужно, обучения не произойдет). A R S Рис. 9. Обозначения A, R и S − как на рисунке 8. Незаполненные круги внутри прямоугольника R − это ON нейроны сетчатки, сигналы от которых в итоге обучения должны проходить на простую клетку, заштрихованные круги вне R ON нейроны, которые имеют синапсы с простой клеткой S, но через эти синапсы в результате обучения сигналы не проходят. Это было о том, как усиливаются синапсы нейронов, соответствующие полоске R, показанной на рисунке 8. Теперь нужно обеспечить, чтобы на простую клетку не воздействовали ON нейроны, находящихся внутри круга A, но вне полоски R. Для этого дальше предполагается, что если простая клетка была возбуждена поступившими на нее сигналами, то находящиеся на ней синапсы ON клеток, через которые не поступало возбуждение, тоже переходят в состояние ожидания; если в промежутке ожидания на простую клетку не поступит сигнала, то с находящимися в состоянии ожидания синапсами ничего не произойдет; если в течение промежутка ожидания на простую клетку поступит возбуждающий сигнал, то синапсы, которые не были возбуждены в предыдущий момент, ослабятся. Таким образом, если на простую клетку поступило возбуждение, то все расположенные на ней синапсы ON нейронов переходят в состояние ожидания. Если в течение промежутка ожидания никакого возбуждения на простую клетку не поступит, то с находящимися на ней синапсами ON нейронов ничего не происходит. Если в промежутке ожидания поступает возбуждающий сигнал, то возбужденные в предшествующий момент синапсы увеличиваются, а синапсы, через которые в предшествующий момент сигнал не поступал, ослабятся. В результате формируется отображение, которое показано на рисунке 9. Но это еще не то отображение, которое было показано на рисунке 1: на рисунке 9 горизонтально заштрихованные круги это не зоны торможения, показанные на 8 рисунке 1, а нейроны, от которых на простую клетку сигнал не поступает. Чтобы получить картину, показанную на рисунке 1, нужно ввести торможение. То есть ввести тормозящие клетки. Исходя из предположения, что ON нейроны находятся в сетчатке, в качестве тормозящих удобно использовать амакриновые клетки. Дальше предполагается, что область, из которой поступают сигналы на простую клетку зрительной коры (рис. 9), находится в слое амакриновых и ганглиозных клеток сетчатки (рис. 10). Из ганглиозных клеток рассматриваются лишь ON клетки. Предполагается, что амакриновые клетки расположены между ними и оказывают на них локальное тормозное воздействие, и что область тормозящего воздействия амакриновой клетки сравнима с радиусом рецептивного поля простой клетки и торможение тем слабее, чем больше расстояние между амакриновой клеткой и ганглиозной клеткой, на которую она воздействует (Cook, Werblin, 1994; Cook et al., 1998; Vaney et al., 1999; Burkhardt, Fahey, 1999; Taylor, 1999; Jacoby, Marshak, 2000; Shields, Lukasiewicz, 2002; Chiao, Masland, 2002; Zhang et al., 2002; Dong, Hare, 2003; Yamada et al., 2003; Kolb, Fernandez, Nelson, 2000/2003). A S Рис. 10. A и S как на рисунке 8. Черные круги амакриновые клетки сетчатки, оказывающие тормозящее воздействие на соседние ганглиозные ON клетки; заштрихованные круги ON нейроны, имеющие синапсы с простой клеткой, но не оказывающие на нее воздействия; незаполненные круги ON нейроны, которые оказывают воздействие на простую клетку. Рассмотрим теперь некоторые особенности функционирования такой структуры. Как было сказано выше, ON нейроны сетчатки, расположенные внутри круга (рис. 10), могут посылать возбуждающий сигнал на синапсы, находящиеся на простой клетке. В результате обучения на простую клетку будут проходить сигналы только от незаштрихованных ON нейронов, а от заштрихованных не будут. Все ON нейроны, находящиеся в радиусе воздействия возбужденных амакриновых клеток, тормозятся этими клетками. Если осветить сетчатку прямолинейной полоской, которая воздействует только на область D (рис. 11), то торможения амакриновыми клетками будет недостаточно, чтобы подавить ганглиозные клетки этой области соответствующая простая клетка S будет возбужденной. Если освещение возбуждает области B или C, а область D освещена слабо (фоновое освещение), то амакриновые клетки будут подавлять ганглиозные клетки области D и сигнал на простую клетку поступать не будет. Картина будет выглядеть так, как будто 9 рецептивное поле простой клетки содержит продолговатую возбуждающую часть и две окаймляющие ее области торможения (рис. 1). A B D S C Рис. 11. Круг A − область сетчатки, внутри которой находятся все ганглиозные ON клетки, посылающие сигнал на простую клетку S, D − область, для которой возбуждающий сигнал от находящихся в ней ганглиозных ON клеток проходит на простую клетку; B и C области, от ON клеток которых сигнал посылается на простую клетку, но на нее не проходит; черные круги тормозящие амакриновые клетки. Из сказанного вытекает, что если тренировать реагировать на более узкую освещенную полоску, например, как на рисунке 12 по сравнению с рисунками 9 и 11, то область, от которой будет поступать сигнал на простую клетку, будет меньшей. В результате возрастет точность настройки простой клетки, но уменьшится число ON нейронов, посылающих сигналы на эту клетку. В результате возбуждение простой клетки и ее ответная реакция будут слабее. Это явление было найдено экспериментально: было установлено, что если тренировать простые клетки более точно отвечать на стимулы определенной ориентации, то интенсивность их выходного сигнала снижается (Schoups et al., 2001; Ghose et al., 2002; Teich, Qian, 2003). 10 A S R Рис. 12. A и S как не рисунке 8; черные круги, заштрихованные круги и незаполненные круги − как на рисунке 10; R прямоугольный стимул, более узкий, чем на рисунке 9. Для простых клеток известно, что с увеличением яркости входного стимула интенсивность их выходного сигнала возрастает. Но при этом настройка этих клеток на ориентацию стимула остается неизменной (рис. 13.a). Это явление называется контрастной инвариантностью (Sclar, Freeman, 1982). В то же время, если исходить из модели Хьюбеля-Визеля, то интенсивность выходного сигнала должна возрастать с ростом интенсивности входного, и тогда для высоких интенсивностей входного сигнала выходной сигнал должен появляться для всех ориентаций стимула (рис. 13.b). 3 2 1 -45 00 a) 450 -45 00 b) 450 Рис. 13. Зависимость интенсивности выходного сигнала простой клетки от ориентации стимула и его интенсивности. Вдоль горизонтальной оси ориентация стимула (в градусах), вдоль вертикальной оси интенсивность выходного сигнала (в условных единицах); a) 1, 2 и 3 примерный вид графиков зависимости интенсивности выходного сигнала простой клетки от входного стимула низкой яркости (1), средней яркости (2) и высокой яркости (3); b) вид тех же графиков, получающийся при использовании модели Хьюбеля и Визеля. Это несоответствие отмечалось многими авторами. Были предложены различные объясняющие его модели, опирающиеся на представления Хьюбеля и 11 Визеля и ряд дополнительных предположений (Troyer et al., 1998; Ferster, Miller, 2000; Troyer et al., 2002; Shapley et al., 2003; Teich, Qian, 2003; Miller, 2003; Ursino, La Cara, 2004). Покажем, что эту инвариантность можно объяснить, если исходить из предположения об обучении простых клеток. A B C Рис. 14. A область ганглиозных ON клеток сетчатки (круг), которые имеют синапсы с данной простой клеткой; C область тех ганглиозных ON клеток, от которых после обучения сигнал проходит на простую клетку (область D на рисунке 11); B область вне C, от которой торможение амакриновыми клетками доходит до ганглиозных ON клеток из C. Рассмотрим область A ганглиозных ON клеток сетчатки, которые имеют синапсы с данной простой клеткой, и область C ганглиозных ON клеток, от которых после обучения сигнал проходит на эту простую клетку. Кроме того, рассмотрим область B вне C, от которой торможение доходит до ганглиозных клеток из C (рис. 14). Поскольку для дальнейшей иллюстрации не понадобятся ганглиозные ON клетки, которые не оказывают влияния на простую клетку, то сохраним только области B и C рисунка 14 (рис. 15). B C Рис. 15. C область, содержащая все ганглиозные ON клетки сетчатки, сигнал от которых после обучения проходит на простую клетку; B (заштриховано) область вне C, из которой торможение амакриновыми клетками достигает ганглиозных ON клеток из C. Осветим сетчатку прямолинейной полоской. Если сигналы от стимула попадут на клетки, расположенные вдоль большой оси области C (рис. 16 а), то возбуждение ганглиозных клеток этой области будет максимальным. Тогда максимальными будут сигналы, поступающие от этой области на простую клетку. Если полоска проектируется под углом к большой оси (рис. 16.b-c), то будет захвачено меньшее число ганглиозных клеток, возрастет вклад торможения амакриновыми клетками и на простую клетку поступит меньшее возбуждение. Из рисунков 16.a-c видно, что при разных углах наклона полоски соотношение интенсивности возбуждения и торможения изменяется. Это значит, что возможно 12 такое соотношение параметров, определяющие возбуждение и торможение, при котором получается связь, показанная на рисунке 13.a. B C a B B C C c b Рис. 16. Те же области возбуждения и торможения, что на рисунке 15, с проекцией на них под разными углами (a-c) возбуждающей прямолинейной полоски. С простыми клетками зрительной коры связана еще одна проблема. Дело в том, что эти клетки образуют примыкающие друг к другу небольшие области, в каждой из которых находятся нейроны с настройкой на одинаковую ориентацию (рис. 17). Рис. 17. Небольшие области разной формы, в которых простые клетки настроены на одинаковую ориентацию предъявляемого зрительного стимула. Существуют разные предположения о возникновении таким образом организованных областей простых клеток (Somers et al., 1995; Ben-Yishai et al., 1995; Shmuel, Grinvald, 2000; Shouval et al., 2000; Ernst et al., 2001; Teich, Qian, 2003; Shapley et al., 2003; Kang et al., 2003; Carriera-Perpiñán, Goodhill, 2004). Ниже рассматривается возможность, которая является следствием описанной выше схемы возникновения простых клеток в результате обучения. Как уже говорилось, на раннем этапе развития организма рецептивное поле простой клетки может быть близким к кругу (рис. 18.a). Если на этом этапе на рецептивное поле попадет изображение прямолинейной полоски, то простая клетка может настроится и отвечать на такую ориентацию отрезка оптимальным образом. Рецептивные поля лежащих рядом простых клеток перекрываются. Это значит, что на появившуюся полоску будут обучаться реагировать несколько клеток (рис. 18.b). В результате возникнет множество расположенных рядом простых клеток, настроенных на одинаково ориентированные полоски. Известно, что глаза новорожденного движутся хаотически. Поэтому воспринимаемая глазом полоска будет последовательно возбуждать некоторое множество рецептивных полей простых клеток. В результате будут возникать поля 13 произвольной формы, состоящие из одинаково ориентированных простых клеток. Если предположить, что полоски различной ориентации появляются в поле зрения независимо, то в результате рядом будут возникать участки простых клеток с разными ориентациями. S S P R R S P S R R a b Рис. 18. Одна простая клетка (a) и группа рядом лежащих простых клеток (b), обучающихся предпочтительно реагировать на полоску данной ориентации, P − полоска данной ориентации, S − простые клетки, R − их рецептивные поля. Заключение. Таким образом, возникновение простых клеток зрительной коры можно легко объяснить, если предположить существование в нервной системе процессов обучения и механизмов, которые оценивают полезность или бесполезность той или иной реакции и соответственно запоминают ее или не запоминают. Представление об обучении объясняет процесс формирования небольших областей одинаково ориентированных простых клеток, а также некоторые особенности их функционирования. При таком подходе одна из основных проблем состоит в возможности проверить, является ли настройка простых клеток на определенную ориентацию врожденной или возникает в результате обучения. Был проведен ряд исследований (Chapman, Stryker, 1993; Chapman et al., 1996; Crair et al., 1998; Sengpiel et al., 1999; Chapman et al., 1999; Dragoi et al., 2000; Krug et al., 2001; Akerman et al., 2004), но их результаты оказались неоднозначными. Такая неоднозначность легко объяснимо. Действительно, чтобы проверить наличие настройки на данную ориентацию, нужно предъявить отрезок с этой ориентацией. Но клетки могут настраиваться на данную ориентацию в процессе проверки, то есть, сама проверка может влиять на эксперимент. (Tavazoie, Reid, 2000; Tian, Copenhagen, 2001, 2003, Xu, Tian, 2004). Таким образом, проблема формирования простых клеток зрительной коры пока остается нерешенной. Как следует из предложенного выше механизма, специальных исследований требует возможность их прижизненного формирования. 14 Литература Adorján, P.; Levitt, J.B.; Lund, J.S.; Obermayer, K. (1999). A model for the intracortical origin of orientation preference and tuning in macaque striate cortex. Visual Neuroscience 16: 303-318. Akerman, C.J.; Grubb, M.S.; Thompson, I.D. (2004). Spatial and Temporal Properties of Visual Responses in the Thalamus of the Developing Ferret. The Journal of Neuroscience 24(1): 170-182. Alonso, J.M.; Usrey, W.M.; Reid, R.C. (2001). Rules of Connectivity between Geniculate cells and Simple Cells in Cat Primary Visual Cortex. The Journal of Neuroscience 21(11): 4002-4015. Ben-Yishai, R.; Bar-Or, R.L; Sompolinsky, H. (1995). Theory of orientation tuning in visual cortex. Proceedings of the National Academy of Science of the United States of America 92: 3844-3848. Burkhardt, D.A.; Fahey, P.K. (1999). Contrast Rectification and Distributed Encoding By ON-OFF Amacrine Cells in the Retina. Journal of Neurophysiology 82: 16761688. Carandini, M.; Ringach, D.L. (1997). Predictions of a recurrent model of orientation selectivity. Vision Research 37: 3061-3071. Carriera-Perpiñán, M.Á.; Goodhill, G.J. (2004). Influence of Lateral Connections on the Structure of Cortical Maps. Journal of Neurophysiology 92: 2947-2959. Casagrande, V.A.; Xu, X.; Sáry, G. (2002). Static and dynamic views of visual cortical organization. In E.S. Azmitia, J. DeFelipe, E.G. Jones, P. Rakic, C.E. Ribak (Eds.): Progress in Brain Research, V. 136, Ch. 30: 389-408. Chapman, B; Gödecke, I. (2000). Cortical Cell Orientation Selectivity Fails to Develop in the Absence of ON-Center Retinal Ganglion Cell Activity. The Journal of Neuroscience 20 (5): 1922-1930. Chapman, B; Gödecke, I.; Bonhoeffer, T. (1999). Development of Orientation Preference in the Mammalian Visual Cortex. Journal of Neurobiology 41: 18-24. Chapman, B; Stryker, M.P.; Bonhoeffer, T. (1996). Development of Orientation Preference Maps in Ferret Primary Visual Cortex. The Journal of Neuroscience 16(20): 6443-6453. Chapman, B.; Stryker, M.P. (1993). Development of orientation selectivity in ferret visual cortex and effects of deprivation. The Journal of Neuroscience 13: 5251-5262. Chiao C.-C.; Masland, R.H. (2002). Starburst Cells Nondirectionally Facilitate the Responses of Direction-Selective Retinal Ganglion Cells. The Journal of Neuroscience 22(24): 10509-10513. Chung, S., Ferster, D. (1998). Strength and orientation tuning of the thalamic input to simple cells revealed by electrically evoked cortical suppression. Neuron 20: 11771189. Cook, P.B.; Lukasiewicz, P.D.; McReinolds, J.S. (1998). Action potentials are required for the lateral transmission of glycinergic transient inhibition in the amphibian retina. The Journal of Neuroscience 18: 2301-2308. Cook, P.B.; Werblin, F.S. (1994). Spike initiation and propagation in wide field transient amacrine cells of the salamander retina. The Journal of Neuroscience 14: 15 3852-3861. Crair, M.C.; Gillespie, D.C.; Stryker, M.P. (1998). The Role of Visual Experience in the Development of Columns in Cat Visual Cortex. Science 279: 566-570. Dong, C.-J.; Hare, W.A. (2003). Temporal Modulation of Scotopic Visual Signals by A17 Amacrine Cells in Mammalian Retina In Vivo. Journal of Neurophysiology 89: 2159-2166. Dragoi, V.; Sharma, J.; Sur, M. (2000). Adaptation-Induced Plasticity of Orientation Tuning in Adult Visual Cortex. Neuron 28: 287-298. Ernst, U.A.; Pawelzik, K.R.; Sahar-Pikielny, C.; Tsodyks, M.V. (2001). Intracortical origin of visual maps. Nature Neuroscience 4(4): 431-436. Ferster, D.; Miller, K.D. (2000). Neural mechanisms of orientation selectivity in the visual cortex. Annual Review of Neuroscience 23: 441-471. Ghose, G.M.; Yang, T.; Maunsell, J.H.R. (2002). Physiological correlates of perceptual learning in monkey V1 and V2. Journal of Neurophysiology 87: 1867-1888. Hebb, D.O. (1949). The organization of behavior. New York. Hubel, D.H.; Wiesel, T.N. (1962). Receptive fields, binocular interaction and functional architecture in the cat's visual cortex. Journal Physiology 160, 106-154. Hubel, D.H.; Wiesel, T.N. (1968). Receptive fields and functional architecture of monkey striate cortex. Journal Physiology 95: 215-243. Jacoby, R.A.; Marshak, D.W. (2000). Synaptic Connections of DB3 Diffuse Bipolar Cell Axons in Macaque Retina. The Journal of Comparative Neurology 416: 19-29. Kang, K.; Shelly, M.; Sompolinsky, H. (2003). Mexican hats and pinwheels in visual cortex. Proceedings of the National Academy of Science of the United States of America 100(5): 2848-2853. Krug, K.; Akerman, C.J.; Thompson, I.D. (2001). Responses of Neurons in Neonatal Cortex and Thalamus to Patterned Visual Stimulation Trough the Naturally Closed Lids. Journal of Neurophysiology 85: 1436-1443. Lund, J.S.; Angelucci, A; Bressloff, P. (2003). Anatomical Substrates for Functional Columns in Macaque Monkey Primary Visual Cortex. Cerebral Cortex 12: 15-24. McLaughlin, D.; Shapley, R.; Shelly, M.; Wielaard, D.J. (2000). A neuronal network model of macaque primary visual cortex (V1): Orientation selectivity and dynamics in the input layer 4Cα. Proceedings of the National Academy of Science of the United States of America 97(14): 8087-8092. Miller, K.D. (1994). A model for the development of simple cell receptive fields and the ordered arrangement of orientation columns through the activity dependent competition between ON- and OFF-center inputs. The Journal of Neuroscience 14: 409-441. Miller, K.D. (2003). Understanding Layer 4 of the Cortical Circuit: A Model Based on Cat V1. Cerebral Cortex 13: 73-82. Reid, R.C.; Alonso, J.M. (1995). Specificity of monosynaptic connections from thalamus to visual cortex. Nature 378: 281-284. Ringach, D.L. (2004). Haphazard Wiring of Simple Receptive Fields and Orientation Columns in Visual Cortex. Journal of Neurophysiology 92: 468-476. Schoups, A.A.; Vogels, R.; Qian, N.; Orban, G.A. (2001). Practising orientation identification improves orientation coding in V1 neurons. Nature 412: 549-553. Sclar, G.; Freeman, R.D. (1982). Orientation selectivity in the cat's striate cortex is 16 invariant with stimulus contrast. Experimental Brain Research, 46: 457-461. Sengpiel, F.; Stawinski, P.; Bonhoeffer, T. (1999). Influence of experience on orientation maps in cat visual cortex. Nature Neuroscience 2: 727-732. Shapley, R.; Hawken, M.; Ringach, D.L. (2003). Dynamics of Orientation Selectivity in the Primary Visual Cortex and the Importance of Cortical Inhibition. Neuron 38: 689-699. Shields, C.R.; Lukasiewicz, P.D. (2002). Spice-Dependent GABA Inputs to Bipolar Cell Axon Terminals Contribute to Lateral Inhibition of Retinal Ganglion Cells. Journal of Neurophysiology 89: 2449-2458. Shmuel, A.; Grinvald, A. (2000). Coexistence 0f linear zones and pinwheels within orientation maps in cat visual cortex. Proceedings of the National Academy of Science of the United States of America 97: 5568-5573. Shouval, H.Z.; Goldberg, D.H.; Jones, J.P.; Beckerman, M.; Cooper, L.N. (2000). Structured Long-Range Connections Can Provide a Scaffold for Orientation Maps. The journal of Neuroscience 20(3): 1119-1128. Smith, E.L.; Chino, Y.M.; Ridder, W.H.; Kitagawa, K.; Langston, A. (1990). Orientation bias of neurons in the lateral geniculate nucleus of macaque monkeys. Visual Neuroscience 5: 525-545. Somers, D.C.; Nelson, S.B.; Sur, M. (1995). An emergent model of orientation selectivity in cat visual cortical simple cells. The Journal of Neuroscience, 15: 54485465. Swindale, N.V. (1996). The development of topography in the visual cortex: a review of models. Network: Computation in Neural Systems 7: 161-247. Tanaka, K (1983). Cross-correlation analysis of geniculostriate neuronal relationship in cats. Journal of Neurophysiology 49: 1303-1318. Tanaka, S. (1991). Theory of ocular dominance column formation. Biological Cybernetics 64: 263-272. Tao, L.; Shelly, M.; McLaughlin, D.; Shapley, R. (2004). An egalitarian network model for the emergence of simple and complex cells in visual cortex. Proceedings of the National Academy of Science of the United States of America 101(1): 366-371. Tavazoie, S.F.; Reid, R.C. (2000). Diverse receptive fields in the lateral geniculate nucleus during thalamocortical development. Nature Neuroscience 3(6): 608-616. Taylor, W.R. (1999). TTX attenuates surround inhibition in rabbit retina ganglion cells. Vision Neuroscience 16: 285-290. Teich, A.F.; Qian, N. (2003). Learning and Adaptation in a Recurrent Model of V1 Orientation Selectivity. Journal of Neurophysiology 98: 2086-2100. Tian, N, Copenhagen, D.R. (2001). Visual deprivation alters development of synaptic function in inner retina after eye opening. Neuron 32: 439-443. Tian, N, Copenhagen, D.R. (2003). Visual stimulation is required for refinement of ON and OFF pathways in postnatal retina. Neuron, 39: 85-96. Troyer, T.W.; Krukowski, A.E.; Miller, K.D. (2002). LGN Input to Simple Cells and Contrast-Invariant Orientation Tuning: An Analysis. Journal of Neurophysiology 87: 2741-2752. Troyer, T.W.; Krukowski, A.E.; Priebe, N.J.; Miller, K.D. (1998). Contrast-invariant orientation tuning in cat visual cortex: thalamocortical input tuning and correlationbased intracortical connectivity. The Journal of Neuroscience 18: 5908-5927. 17 Ursino, M.; La Cara, G-E. (2004). Comparison of different models of orientation selectivity based on distinct intracortical inhibition rules. Vision Research 44: 16411658. Usrey, W.M.; Alonso, J.-M.; Reid, R.C. (2000). Synaptic Interactions between Thalamic Inputs to Simple Cells in Cat Visual Cortex. The Journal of Neuroscience 20(14): 5461-5467. Vaney, D.I.; Nelson, J.C.; Pow, D.V. (1998). Neurotransmitter Coupling through Gap Junctions in the Retina. The Journal of Neuroscience, 18(24): 10594-10602. Vidyasagar, T.R.; Pei, X.; Volgushev, M. (1996). Multiple mechanisms underlying the orientation selectivity of visual cortical neurons. Trends in Neuroscience 19: 272277. Von der Malsburg, C. (1973). Self-organization of orientation sensitive cells in the striate cortex. Kybernetik 14: 85-100. Wielaard, D.J.; Shelly, M.; McLaughlin, D.; Shapley, R. (2001). How Simple Cells Are Made in a Nonlinear Network Model of the Visual Cortex. The Journal of Neuroscience 21(14): 5203-5211. Wong, R.O.L. (1999). Retinal waves and visual system development. Annual Review of Neuroscience 22: 29-47. Xu, H., Tian, N. (2004). Pathway-Specific Maturation, Visual Deprivation, and Development of Retinal Pathway. Neuroscientist 10(4): 337-346. Yamada, E.S.; Dmitrieva, N.; Keyser, K.T.; Lindstrom, J.M.; Hersh, L.B.; Marshak, D.W. (2003). Synaptic Connections of Starburst Amacrine Cells and Localization of Acetilcholine Receptors in Primate Retinas. The Journal of Comparative Neurology 461: 76-90. Zhang, J.; Li, W.; Trexler, E.B.; Massey, S.C. (2002). Confocal Analysis of Reciprocal Feedback at Rod Bipolar Terminals in the Rabbit Retina. The Journal of Neuroscience 22(24): 10871-10882. Zhou, Y.; Leventhal, A.G.; Thompson, K.J. (1995). Visual deprivation does not affect the orientation and direction sensitivity of relay cells in the lateral geniculate nucleus of the cat. Journal of Neuroscience 15: 689-698.