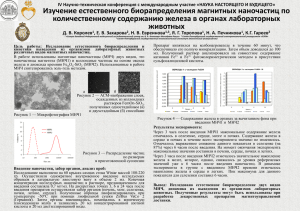
адаптивные реакции и пределы толерантности triticum aestivum
реклама

ФЕДЕРАЛЬНОЕ ГОСУДАРСТВЕННОЕ БЮДЖЕТНОЕ НАУЧНОЕ УЧРЕЖДЕНИЕ ВСЕРОССИЙСКИЙ НАУЧНО-ИССЛЕДОВАТЕЛЬСКИЙ ИНСТИТУТ МЯСНОГО СКОТОВОДСТВА На правах рукописи ДЕРЯБИНА ТАТЬЯНА ДМИТРИЕВНА АДАПТИВНЫЕ РЕАКЦИИ И ПРЕДЕЛЫ ТОЛЕРАНТНОСТИ TRITICUM AESTIVUM И ALLIUM CEPA К НАНОЧАСТИЦАМ МЕДИ И ЖЕЛЕЗА 03.02.01 – ботаника 03.02.08 – экология Диссертация на соискание ученой степени кандидата биологических наук Научный руководитель: доктор биологических наук, профессор С.А. Мирошников ОРЕНБУРГ – 2015 2 СОДЕРЖАНИЕ ВВЕДЕНИЕ………………………………………………..…………………………....4 ГЛАВА 1. ДИСПЕРСИИ МЕТАЛЛОВ В ЕСТЕСТВЕННЫХ И ТЕХНОГЕННО-ИЗМЕНЕННЫХ ЭКОСИСТЕМАХ. РЕАКЦИЯ РАСТЕНИЙ И МИКРООРГАНИЗМОВ НА ВОЗДЕЙСТВИЕ НАНОЧАСТИЦ МЕТАЛЛОВ (ОБЗОР ЛИТЕРАТУРЫ)………………………………………………………………………….9 1.1 Адаптация растений к условиям среды обитания с различным содержанием минеральных веществ…………………………………………….………………...….9 1.2 Общие аспекты взаимодействия высокодисперсных веществ окружающей среды и растений……………………………………………………………...………17 1.3 Приспособление растений к воздействию техногенных наноматериалов. Взаимодействие в системе «наночастицы – растения»…………………………….21 1.4. Резюме…………………………………………………………………………….36 ГЛАВА 2. МАТЕРИАЛЫ И МЕТОДЫ ИССЛЕДОВАНИЯ……………….………......38 2.1 Объекты исследования………………………………………….…………...……38 2.1.1. Растительные и бактериальные тест – объекты…………………..…………..38 2.1.2. Наночастицы и другие соединения меди и железа…………………………..38 2.1.3. Наночастицы и другие соединения меди железа………….............................39 2.2 Методы исследования…………………………………………………………….41 2.2.1 Методы исследования с использованием растительных организмов……….41 2.2.1.1 Культивирование растительных организмов в контакте с наночастицами меди и железа с оценкой их толерантности к подобному воздействию….……….41 2.2.1.2 Методы микроскопического исследования растительных организмов…...45 2.2.1.3 Методы оценки накопления и распределения меди и железа в тканях растительных организмов…………………………………………………………….46 2.2.1.4 Метод анализа ДНК растительных организмов…………………………….48 2.2.2 Методы исследования с использованием люминесцирующих бактерий…...49 2.2.2.1 Метод оценки токсичности соединений меди и железа в тесте тушения бактериальной биолюминесценции………………………………………………….49 2.2.2.2 Метод оценки механизмов биоактивности соединений меди и железа 3 в тесте индукции бактериальной биолюминесценции………………………..……51 2.2.3 Статистическая обработка результатов……………………………………….52 ГЛАВА 3. СРАВНИТЕЛЬНАЯ ОЦЕНКА ТОЛЕРАНТНОСТИ РАСТЕНИЙ (В ТЕСТЕ ПРОРАСТАНИЯ СЕМЯН TRITICUM AESTIVUM) И МИКРООРГАНИЗМОВ (ТЕСТЕ ИНГИБИРОВАНИЯ БИОЛЮМИНЕСЦЕНЦИИ SHERICHIA COLI) ОЗДЕЙСТВИЮ НАНОЧАСТИЦ, МИКРОЧАСТИЦ И СОЛЕЙ ЖЕЛЕЗА И МЕДИ……….……………………………………………………………53 3.1. Оценка безопасности ионов, нано- и микрочастиц Fe и Cu в тесте прорастания семян Triticum aestivum………………………………..………………………….…..53 3.2. Биологическая активность ионов, нано- и микрочастиц Cu и Fe в тесте ингибирования бактериальной биолюминесценции………………………………..57 ГЛАВА 4. АДАПТИВНЫЕ РЕАКЦИИ РАСТЕНИЙ К ПРИСУТСТВИЮ В СРЕДЕ КУЛЬТИВИРОВАНИЯ НАНОЧАСТИЦ МЕДИ…………………………………...63 4.1. Анализ механизмов биологической активности наночастиц меди в тестах на Triticum aestivum и Allium cepa……………………………………………………….63 4.2. Исследование механизмов ДНК-повреждающего эффекта наночастиц меди в тестах на люминесцирующих штаммах Escherichia coli…………………………...75 ГЛАВА 5. АДАПТИВНЫЕ РЕАКЦИИ TRITICUM AESTIVUM К ПРИСУТСТВИЮ В СРЕДЕ КУЛЬТИВИРОВАНИЯ НАНОЧАСТИЦ ЖЕЛЕЗА..86 ЗАКЛЮЧЕНИЕ………………………………………………………………………..92 ВЫВОДЫ…………………………………………...………………………………..104 СПИСОК ИСПОЛЬЗОВАННОЙ ЛИТЕРАТУРЫ…………………………………106 4 Введение Актуальность темы. Дисперсные системы, содержащие минеральные наночастицы, являются естественным компонентом большинства природных экосистем. О типичности подобной ситуации свидетельствуют данные, согласно которым 1 м³ атмосферного воздуха содержит до 1 млн взвешенных частиц диаметром от 0,001 до 1000 мкм, их концентрация в пресной и морской воде достигает 5-6 млн/дм³, а наиболее существенно дисперсные системы представлены в почве (Yushkin N.P. Mineralis mundi et biosphere: mineralisorganizmobioz, biomineral interaction coevolution // Acta IV International seminar «Mineralogy et vitam: Origin de biosphere et co-evolution of mineralis et biologicummundosbiomineral-ogiya». Syktyvkar. 2007. P. 5-7). Кроме того, ожидаемый рост производства и использования наночастиц создает дополнительные риски их поступления в антропогенно-измененные экосистемы (Gottschalk F., Sonderer T., Scholz R.W. Modeled environmental concentrations of engineering nanoparticles (TiO2, ZnO, Ag, CNT, fullerenes) for different regions // Environ. Sci. Technol. 2009. Vol.43. P. 9216 – 9222). Однако, с позиций факториальной экологии, изучающей воздействие факторов окружающей среды на живые организмы, наночастицы оказываются еще относительно слабо охарактеризованными объектами, интерес к исследованию которых в ближайшее время будет только возрастать. Таким образом, складывается ситуация, свидетельствующая об актуальности экологической аттестации наночастиц металлов, в том числе закономерностей их поступления, распределения и накопления в компонентах биоты, а также выявления разнообразия обусловленных этим биологических эффектов и механизмов их формирования. Степень разработанности темы. Среди живых систем, потенциально подвергающихся воздействию наночастиц, важное место занимают растения, традиционно в используемые различных системах биоиндикации и биотестирования (Kahru A., Dubourguier H., Blinova I. et al. Biotests and biosensors for ecotoxicology of metal oxide nanoparticles: a minireview // Sensors. 2008. Vol. 8. P. 5153-5170). При этом особый интерес к растениям обусловлен их базовым 5 положением в структуре пищевых цепей, а также позитивным опытом использования в системе биологической ремедиации, что в обоих случаях связано со способностью к накоплению разнообразных химических элементов. Адекватность подобного подхода для оценки биологической активности наночастиц и наноматериалов находит подтверждение и в ряде современных исследований (Lin D. Phytotoxicity of nanoparticles: inhibition of seed germination and root growth // Environmental Pollutants. 2007. Vol. 150. Iss. 2. P. 243-250; Musante C., White J.C. Toxicity of silver and copper to Cucurbitapepo: differential effects of nano and bulk-size particles // Environ. Toxicol. 2012. Vol. 27. P. 510-517), позволивших оценить характер воздействия наночастиц ряда металлов и оксидов металлов на рост и развитие модельных растений. В Российской Федерации подобный подход закреплен в МУ 1.2. 2635-10 «Медико-биологическая оценка безопасности наноматериалов» (2010), а также в ряде других нормативнометодических документов. Между тем, задача оценки толерантности растений к действию наночастиц металлов, равно как и изучение механизмов их адаптации к присутствию подобных наночастиц в среде обитания, пока полностью не решены и требуют своего дальнейшего исследования. Целью работы явилось изучение адаптивных реакций и пределов толерантности Triticum aestivum L. и Allium cepa L. к условиям среды, содержащей наночастицы меди и железа, а также оценка механизмов биологической активности данных наночастиц в отношении растительных организмов. Для достижения обозначенной цели были поставлены следующие задачи: 1. Дать сравнительную оценку толерантности растений к наночастицам Cu и Fe в тестах прорастания семян Triticum aestivum в сравнении с чувствительностью к ионам и микрочастицам этих металлов. 2. Изучить особенности роста и развития растений (на моделях Triticum aestivum и Allium cepa) при культивировании в среде с наночастицами Cu с идентификацией механизма их действия в отношении живых систем. 3. Охарактеризовать особенности адаптации Triticum aestivum условиям среды, содержащей субтоксические концентрации наночастиц Fe. к 6 Научная новизна работы состоит в том, что в ней впервые определены особенности роста и развития растений Triticum aestivum и Allium cepa при культивировании в среде содержащей нано-, микрочастицы и ионы железа или меди. Для организмов различного уровня организации показано согласованное изменение значений верхнего предела толерантности, прогрессивно убывающего в ряду «микрочастицы → наночастицы → ионы», толерантность к соединениям а также более высокая железа в сравнении с соединениями меди. Установлено, что растения преимущественно реагируют на подобное воздействие нарушением образования корневой системы. Показана возможность поступления наночастиц меди из среды культивирования в ткани корня растений через систему межклеточных контактов. Зафиксировано формирование окислительного стресса в клетках, взаимодействующих с наночастицами меди, следствием которого является повреждение присутствующей в них ДНК. Представлены данные о деградации ДНК и подавлении митотической активности в корневой системе растений при их проращивании в контакте с наночастицами меди, объясняющем природу развивающегося фитотоксического эффекта. Продемонстрирован умеренно выраженный стимулирующий эффект субтоксических концентраций наночастиц железа на рост и развитие Triticum aestivum. Теоретическая и практическая значимость работы состоит в развитии и совершенствовании методического аппарата для анализа биологической активности вновь синтезируемых наночастиц и наноматериалов с обоснованием оптимального алгоритма проведения подобных исследований, в том числе с использованием взаимодополняющих биотестов. Полученные результаты могут быть использованы в системе обоснования допустимых уровней загрязнения растительных экосистем и агроценозов, существенным компонентом которых являются металлические дисперсии. Ростостимулирующий эффект субтоксических концентраций некоторых наночастиц расширяет показания к их использованию при предпосевной обработке семян, а также при выращивании кормовых и гидропоники. сельскохозяйственных растений методами аэропоники или 7 Методология и методы исследования. При проведении исследований использованы дополняющие базовые их микроорганизмов, подходы биотесты применение экспериментальной с ботаники, использованием которых при а также люминесцирующих исследовании биологической активности наночастиц одобрено действующими национальными нормативами. Спектр конкретных методов включал тест прорастания семян Triticum aestivum, классический и модифицированный варианты Allium-теста, микроскопические и электронно-микроскопические наблюдения, метод атомно-адсорбционной спектрометрии, бактериальные биотесты для оценки окислительного стресса, а также генетические тесты оценки повреждения ДНК. Все эксперименты выполнены в нескольких повторностях, а полученный фактический материал обработан с использованием методов статистического анализа. Положения, выносимые на защиту: 1. Толерантность растений к наночастицам металлов превосходит таковую к их ионным формам, но уступает микрочастицам, при этом данные организмы более толерантны к соединениям железа, нежели к соединениям меди. 2. Низкая адаптивная способность растений Triticum aestivum и Allium cepa к условиям среды обитания со значительным содержаниями наночастиц меди может быть связана с повреждением ДНК растительных клеток, следствием чего является митотоксический эффект (подавление размножения клеток до вхождения в митоз), а его итоговым проявлением – фитотоксический эффект, заключающийся в нарушении роста корневой системы растений. 3. Включение в среду культивирования наночастиц железа в определенном диапазоне концентраций стимулирует рост Triticum aestivum . Личный вклад автора. Автором самостоятельно осуществлена постановка цели и определены основные задачи диссертационной работы, выбраны и обоснованы необходимые для их достижения методы исследований. Анализ присутствия металлов в тканях модельных растений проведен совместно с сотрудниками ЦКП «Институт микро- и нанотехнологий» ОГУ. Электронномикроскопические исследования выполнены на базе ЦКП «Симбиоз». Автором 8 лично выполнена основная часть экспериментов, проведена их математическая обработка, анализ и обобщение полученных результатов. Подготовка к печати научных работ, отражающих результаты диссертации, осуществлялась самостоятельно или при участии соавторов. Основной блок работ проводился в 2011-2014 годах в период обучения в очной аспирантуре и работе в ФГБНУ ВНИИМС. Организация исследований. Отдельные фрагменты работы выполнены при финансовой поддержке Государственного задания на проведение научных исследований по теме «Исследование прямых и отдаленных эффектов влияния наночастиц металлов на биологические системы, обеспечивающие разработку функциональных трофических субстратов» (рег. № 4.2979.2011 / 1.8.11), а также гранта РНФ «Исследование по проблемам агроэкологии техногенных наноматериалов, обеспечивающих улучшение условий жизни и среды обитания человека» (проект №14-36-00023). Степень достоверности и апробация работы. Научные положения и выводы обоснованы и базируются на воспроизводимых экспериментальных данных, степень достоверности которых доказана путем статистической обработки. Основные положения диссертационной работы были доложены и обсуждены на V и VI Всероссийских научно-практических конференциях «Проблемы экологии Южного Урала» (Оренбург, 2011, 2013); III съезде Российского общества медицинской элементологии (Москва, 2012). Публикации. По материалам диссертации опубликовано 7 работ, в том числе 6 статей в рецензируемых журналах, рекомендуемых ВАК РФ для публикации результатов диссертационных исследований, получен 1 патент РФ на изобретение. Объем и структура диссертации. Диссертация состоит из введения, 5 глав, заключения и выводов, включает 123 страницы, 9 таблиц и 14 рисунков. Список литературы представлен 175 наименованиями, из них 71 на иностранных языках. 9 ОБЗОР ЛИТЕРАТУРЫ ГЛАВА 1. ДИСПЕРСИИ МЕТАЛЛОВ В ЕСТЕСТВЕННЫХ И ТЕХНОГЕННО-ИЗМЕНЕННЫХ ЭКОСИСТЕМАХ. РЕАКЦИЯ РАСТЕНИЙ НА ВОЗДЕЙСТВИЕ НАНОЧАСТИЦ МЕТАЛЛОВ 1.1 Адаптация растений к условиям среды обитания с различным содержанием минеральных веществ Одним из важнейших качеств растений является способность адаптироваться к меняющимся условиям окружающей среды, что обеспечивает их выживание, устойчивость к воздействию факторов абиотического и биологического характера, успех в конкуренции с другими видами. С общебиологической точки зрения адаптация (от латинского аdaptation – приспособление) может быть условно разделена на генотипическую и фенотипическую (Илющенко В.Г. Классификация спонтанной генотипической клеточной адаптации // Цитология и генетика. 2002. Т.36 №5. С. 34-42). Следует отметить, что эти два начала тесно переплетены между собой и определяют перспективы выживания видов в долгосрочной и краткосрочной перспективе. При этом адаптация, как правило, реализуется последовательно от изменений во внешней среде растения к изменению строения и функции. Наукой накоплен огромный багаж знаний о структурных и функциональных изменениях в растительных организмах при адаптации. В частности, адаптация растения земляники к недостатку влаги в почве сопровождается снижением толщины листовой пластинки вследствие мелкоклеточности – ксероморфизма (Кушниренко М.Д. Физиология водоомена и засухоустойчивости плодовых растений. Кишинев, 1975); водоудерживающая способность клеточных коллоидов листьев повышается, ее содержание в растениях растет (Хвостов Д.С. Морфоанатомические и физиологические механизмы приспособления растений земляники к сезонным изменениям условий внешней среды // Сельскохозяйственная биология. 2004. №5. С.89-93). Адаптация J. communis var. 10 saxatilis к высокогорью и низким температурам сопровождается развитием мезофилла, формированию крупных смоленных каналов хвои (Князева С.Г. Морфолого-анатомические особенности хвои можжевельника обыкновенного (juniperus communis L.) // Хвойные бореальной зоны. 2012. ХХХ. №1-2. С. 92-96). Для растений H. gmelinii адаптация к высокогорью сопряжена с изменениями морфоструктуры генеративных побегов (Горышина Т.К. Экология растений. М.: Высшая школа, 1979. 368 с.) и угнетением интеркалярного роста меристем (Жмудь Е.В. Экологическая пластичность Hegysarum Gmtlinii (fabactae) в Горном Алтае и Хакасии // Вестник Томского Государственного политехнического университета. 2014. №11(152). С. 220-226). Очевидно, что на протяжении всей истории высших растений адаптация к трофическим факторам оказывала решающее влияние на их эволюцию, способствуя появлению или гибели отдельных видов, изменяя признаки существующих генераций. Это в полной мере относится и к такой важной составляющей этой группы факторов как минеральное питание (Veen B.W., Kleinendorst A. The role of nitrate in osmoregulation of Italian ryegrass // Plant and Soil. 1986. V.91. P.433-436; Трапезников В.К, Иванов И.И., Тельвинская Н.Г. Локальное питание растений. Уфа: Гилем, 1999. 260 с.; Черкозьянова А.В. Координация роста побега и корня проростков пшеницы в условиях дефицита минерального питания: дис. канд. биол. наук. Уфа, 2005. 139 с). Рост и развитие, равно как и адаптация растений невозможны без шестнадцати основных химических элементов (Glass A.D.M. Regulation of ion transport // Annu. Rev. Plant Physiol. -1983.- V.34. - P.311-326). Однако, существуют отдельные виды растений, жизнедеятельность которых невозможна без специфических элементов (Черкозьянова А.В. Координация роста побега и корня проростков пшеницы в условиях дефицита минерального питания: дис. канд. биол. наук. Уфа, 2005. 159 с.). Селен жизненно важен для морской водоросли Thalassiosira psendonana (Price N.M., Thomson P.A., Harrison P.J. Selenium: An essential element for growth of the coastal marine diatom Thalassiosira pseudonana // J. Phyol. V.23. P.1-9); натрий - 11 для Atriplex vesicaria; кремний - для диатомовых водорослей Eguisetum arvense и др. (Werner D., Roth R. Silica metabolism // Inorganic Plant Nutrition. Encyclopedia of Plant Physiology, New Series / Eds. A. Lauchli, R.L. Bieleski. Berlin: Spring-Verlag, 1983. V.15B. P.682-694). В тоже время жизненная необходимость химических элементов для растений определяется их физиологическими функциями. Так, поступление азота, как составной части белка, регламентирует скорость и характер физиологических процессов в растениях, влияет на органообразование, количество и качество сухого вещества (Сабинин Д.А. Избранные труды по минеральному питанию. М.: Наука, 1971. 512 с; Трапезников В.К, Иванов И.И., Тельвинская Н.Г. Локальное питание растений. Уфа: Гилем. 1999. 260 с.). В свою очередь, калий необходим для построения и «работы» более чем 50 ферментов (Evens H.J. Wildes R.A. Potassium and its role in enzyme activation // Potassium in Biochemistry and Physiology. Proc.8 Colloq. Int. Potash Inst. Bern. 1971. P.13-39); обеспечивает особую ионную среду для белкового синтеза (Leigh R.A., Wyn Jones R.G. An hypothesis relating critical potassium concentrations for growth to the distribution and functions of this ion in the plant cell // New Phytologist. 1984. V.97. P.1-13); нормализует осмотическое давление (Haschke K.P., Luttge K. Interactions between IAA, potassium and malate accumulation and growth in Avena coleopite segments // Z. Pflanzenphysiol. 1975. V.76. P.450-455); необходим для открытия устьиц и для многих двигательных процессов растений (Satter R.L., Applewhite P.B., Galston A.W. Rhythmic potassium flux in Allbizzia. Effect of aminophylline, cations and inhibitors of respiration and protein synthesis // Plant Physiology. V.54. P.280-285). В виду того, что концентрация доступных химических элементов в почвах может сильно варьировать, высшие растения в ходе эволюции приобрели целый ряд адаптационных свойств, включающих механизмы аккумуляции, удаления и компартментации ионов, обеспечивающих создание уникальной химической среды, в которой могут протекать обменные процессы (Glass A.D.M. Regulation of ion transport // Annu. Rev. Plant Physiol. 1983. V.34. P.316). 12 Адаптация растительных клеток к условиям среды с избыточным содержанием химических элементов в пределах толерантной зоны (нормы реакции), сопровождается изменением антиоксидантного статуса, активности ферментов (АТФ-аз, фосфатаз, гидролаз, липаз и др.), изменением жирнокислотного состава мембран, направленности энергетических процессов (Mehrle P.M., Bergmann H.L. Biomarkers: Biochemical, Physiological, Histological Markers of Anthropogenic Stress. Lewis: Boca Raton, FL, USA, 2002. Р. 211–234). При хронических токсичных влияниях и высоких концентрациях веществ, когда исчерпываются базовые защитные ресурсы в клетке, происходит спонтанное протекание неспецифических явлений: неконтролируемая проницаемость мембран и деполяризация мембранного потенциала плазмалеммы (Веселова Т.В. Веселовский В.А., Чернавский Д.С. Стресс у растений (Биофизический подход). М.: РАН, 1993. 144 с.), увеличение концентрации ионов кальция в цитоплазме, закисление цитоплазмы, активация микрофиламентов (Пахомова В.М. Основные положения современной теории стресса и неспецифический адаптационный синдром у растений // Цитология. 1995. Т. 37, Вып. 1-2. С. 66–87), увеличение затрат АТФ, хаотическое развитие свободнорадикальных процессов, усиление активности протонного насоса в плазмалемме с одновременным разобщением окисления и фосфорилирования и как следствие структурно-функциональные модификации белков и др. (Чиркова Т.В. Клеточные мембраны и устойчивость растений к стрессовым воздействиям // Соросовский образовательный журнал. 1997. № 9. С. 12–17; Грубинко В.В., Костюк К.В. Структурные адаптации клеточных стенок водных растений к действию цинка и свинца // Бiологiчний вiсник МДПУ. 2012. №2. С. 58-66). Хорошо известны механизмы адаптации, определяющие выживание и процветание отдельных видов растений в условиях засоленности (Строгонов Б.П. Физиологические основы солеустойчивости растений. М.: Изд. АН СССР. 1962. С. 366; Захарин А.А. Особенности водно-солевого обмена растений при солевом стрессе // Агрохимия. 1990. №8. С. 69-79; Керимов Ф.А., Кузнецов Вл.В., Шамина З.Б. Организменный и клеточный уровни солеустойчивости двух сортов 13 хлопчатника [133, ИНЭБР-85] // Физиология растений. 1993. Т.40. №1. С. 128-131; Удовенко Г.В. Принципы различной реакции сортов и видов растений на засоление почвы // Сорт и удобрение. Иркутск. 1974. С. 219-223; Шахов А.А. Солеустойчивость растений. - М.: Изд-во АН СССР. 1956. С. 552; Greenway H., Rana Munns. Mechanisms of salt tolerance in nonhalophytes // Annual Review of Plant Physiology. 1980. V.31. P. 149-190). В частности, одним из механизмов устойчивости растений к значительному содержанию в среде обитания солей является накопление в тканях низкомолекулярных соединений пролина (Stewart G.R., Lee J. A. The rate of Proline Accumulation in Halophytes // Planta Berl. 1974. Vol.120. P. 279-289; Бритиков Е.А. Биологическая роль пролина. М.: Наука. 1975. С. 88) Пролин выполняет в растениях в условиях стресса роль осморегулятора или протектора (Шевякова Н.И., Каролевски П. К вопросу о механизмах ответных реакций на засоление различных по солеустойчивости сортов фасоли // Сельскохозяйственная биология. 1994. №1. С. 84-88). Универсальность систем гомеостатирования внутренней среды позволила высшим растениям закрепиться в своей экологической нише и привела к их сегодняшнему процветанию. Между тем окружающий нас мир претерпевает непрерывные изменения, и существующие виды растений испытывают влияние целого ряда антропогенных факторов. К числу последних можно отнести значительное поступление в среду тяжелых металлов. Накопление тяжелых металлов в клетках и тканях растений сопровождается структурно-функциональными нарушениями, что отрицательно влияет на физиологические процессы, в отдельных случаях ведет к их гибели (Prasad M. N. V. Cadmium toxicity and tolerancein vascular plants // Environ. Exp. Bot. 1995. Vol. 35. P. 525–545; Sanità di Toppi L., Gabbrielli R. Response to cadmium in higher plants // Environ. Exp. Bot. 1999. Vol. 41. P. 105–130; Серегин И. В., Иванов В. Б. Физиологические аспекты токсического действия кадмия и свинца на высшие растения // Физиология растений. 2001. Т. 48, № 4. С. 606–630; Clemens S. Toxic metal accumulation, responses to exposure and mechanisms of tolerance in plants // 14 Biochimie. 2006. Vol. 88. P. 1707–1719; Титов А. Ф., Таланова В. В., Казнина Н. М., Лайдинен Г. Ф. Устойчивость растений к тяжелым металлам. Петрозаводск: Карельский НЦ РАН. 2007. 170 с; Wahid A., Arshad M., Farooq M. Cadmium phytotoxicity: responses, mechanisms and mitigation strategies // Organic farming, pest control and remediation of soil pollutants / Ed. E. Lichtfouse. Springer Science+Business Media B.V. 2009. P. 371–403). Показано также, что скорость накопления, а также характер распределения тяжелых металлов по органам растений зависят от условий выращивания и биологических особенностей вида (Baker A. J. M., Walker P. M. Ecophysiology of metal uptake by tolerant plants // Heavy metal tolerance in plants: evolutionary aspects. Boca Raton: CRC Press,1990. P. 79-87.; Ильин В. Б. Тяжелые металлы в системе почва – растение. Новосибирск: Наука, 1991. 150 с.; Wagner G. J. Accumulation of cadmium in crop plants and consequences to human health // Advances in Agronomy. 1993. Vol. 51. P. 173–212.; Башмаков Д. И., Лукаткин А. С. Эколого-физиологические аспекты аккумуляции и распределения тяжелых металлов у высших растений. Саранск: Изд-во Мордов. Ун-та, 2009. 236 с.) Ввиду того, что поступление ионов тяжелых металлов в растения осуществляется с помощью ионтранспортных систем (транспортных белков, ионных каналов), обеспечивающих поглощение эссенциальных элементов (Hall J. L., Williams L. E. Transition metal transporters in plants // Journal of Experimental Botany. 2003. Vol. 54, N 393. P. 2601–2613; Clemens S. Toxic metal accumulation, responses to exposure and mechanisms of tolerance in plants // Biochimie. 2006. Vol. 88. P. 1707–1719; Krämer U., Talke I. N., Hanikene M. Transition metal transport // FEBS Letters. 2007. Vol. 581. P. 2263–2272), то и механизмы адаптации растений к их избыточному поступлению сходны. Для растения, как целого организма, эти механизмы могут выражаться снижением поступления веществ из корневой системы в стебель и листья. Так, корневой барьер играет заметную роль в формировании толерантности к избытку меди (Демедчик В.В., Соколик А.И., Юрин В.М. Токсичность избытка меди и толерантность к нему растений // Успехи современной биологии, 2001. №5. Т.121. С. 511-525). Подобная картина 15 наблюдалась И.Х. Гаджиевой и др., (2010) в отношении ионов цинка в эксперименте на модели проростков огурца (Гаджиева И.Х., Алиева З.М., Рамазанова П.Б. Кросс-адаптация растений к почвенному засолению и тяжелым металлам // Юг России: экология, развитие. 2010. №1. С. 26-32). В условиях лабораторного опыта (Батова Ю. В., Титов А. Ф., Казнина Н. М., Лайдинен Г. Ф. Накопления кадмия и его распределению по органам у растений ячменя разного возраста // Труды Карельского научного центра РАН. 2012. №2. С. 32–37) по оценке накопление кадмия и его распределение по органам у растений ярового ячменя (Hordeum vulgare L.) установлено, что содержание кадмия в органах с увеличением возраста растений возрастает в корнях и стеблях, в то время как в листьях не изменяется или даже несколько снижается. В тоже время адаптация растений к избытку тяжелых металлов в среде распространяется и на клеточные и внутриклеточные процессы. Все эти механизмы, в конечном итоге, определяют приспособление растений к условиям внешней среды. В этой связи, определенный интерес представляют механизмы адаптации растений к новым классам веществ – техногенным наноматериалам, которые, будучи синтезированными человеком получают все большее распространение в биосфере. Не смотря на то, что развитие нанотехнологий (НТ) связано с началом 21 века, история этого научного направления охватывает не одно столетие. Основы нанотехнологии, как таковой, были заложены во второй половине XIX века в связи с развитием коллоидной химии. В 1857 году М. Фарадей впервые получил устойчивые коллоидные растворы (золи) золота, имеющие красный цвет. В 1861 году Т. Грэму удалось провести коагуляцию золей и превратить их в гели. Он также ввел деление веществ по степени дисперсности структуры на коллоидные (аморфные) и кристаллоидные (кристаллические). В 1869 году химик И. Борщов высказал гипотезу, что вещество, в зависимости от условий, может быть получено и в кристалловидном (склонность к образованию кристаллов), и в коллоидном (аморфном) состоянии (Врублевский Э., Киреев В., 16 Недзвецкий В., Сосновцев В. Нанотехнология – путь в будущее или бренд для финансирования // Нано- и микросистемная техника. 2007. № 12. С. 6–20). Последние два десятилетия нанотехнологии получили значительное развитие, что было обусловлено необходимостью совершенствования имеющейся материальной базы современных электронных устройств. Именно во многом благодаря этому перечень наноматериалов, выпускаемых промышленностью, превысил 1800 наименований, но это число и область их использования постоянно расширяются (Кисель В.П. Нетрадиционные природные ресурсы, инновационные технологии и продукты // Сб. науч. тр. РАЕН. М., 2003. Вып. 10. С. 183-196). Наноматериалы нашли свое применение и в растениеводстве. Так, известно применение наночастиц эссенциальных металлов для стимулирования и защиты семян растений (Крылов С.В., Паничкин Л.А., Захарин А.А. и др. Способ оценки метода протравливания семян // Патент РФ №2015631 от 15.07.1994 г.; Крылов С.В. и др. Состав для обеззараживания семян // Патент РФ №2074616 от 10.03.1997 г.); в качестве микроудобрений (Райкова А.П., Паничкин Л.А., Райкова Н.Н. Исследования влияния ультрадисперсных порошков металлов, полученные различными способами на рост и развитие растений // Материалы междунар. науч.-техн. конф. «Нанотехнологии и информационные технологии – технологии 21 века».- М., 2006. С. 118-123.) и т.д. Между тем, перечень наноматериалов столь обширен и столь быстро увеличивается, что уже в ближайшем будущем растения, в частности, экосистемы, в общем, будут подвержены их воздействию. Однако, несмотря на то, что наноматериалы получают все большее распространение, ни один вид наноматериалов не был изучен в полном объеме по влиянию на биосферу. Фактически во всем мире проводилось незначительное количество таких исследований, которые не позволяют точно оценить потенциальные риски использования наноматериалов (Кисель В.П. Нетрадиционные природные ресурсы, инновационные технологии и продукты // Сб. науч. тр. РАЕН. М., 2003. Вып. 10. С. 183-196). 17 В тоже время результаты исследований большой группы экспериментаторов (Hao, L., Wang, Z // J.Environ.Sci. (Китай). 2009. V.21. P.14591466; Hillegass J.M., Shukla J. M., Lathrop S. // Wiley. Interdiscip. Rev. Nanomed. Nanobiotechnol. 2010. V.2. P. 219-231) и др., наглядно демонстрируют наличие токсических эффектов со стороны наноматериалов на живые организмы. Многочисленные исследования обнаружили прямую связь уровня загрязненности воздуха наночастицами техногенной (угольная, резиновая пыль) и бытовой (табачный дым, пыль) природы – вблизи автотрасс, теплостанций и заводов – с состоянием растительных сообществ. Токсичность наноматериалов, согласно имеющимся литературным данным, может быть обусловлена, развитием окислительного стресса, что приводит к развитию воспалительной реакции, апоптозу и некрозу клетки. Нельзя исключать, однако, и наличия других механизмов токсичности наноматериалов, связанных, в частности, с их повреждающим действием на клеточные мембраны и органеллы, усилением транспорта потенциально токсичных компонентов через барьеры организма, а также возможной генотоксичностью (Глушкова А.В., Радилов А.С., Рембовский В.Р. Нанотехнологии и нанотоксикология – взгляд на проблему // Методологические проблемы изучения и оценки био- и нанотехнологий (нановолны, частицы, структуры, процессы, биообъекты) в экологии человека и гигиене окружающей среды: материалы пленума Научного совета по экологии человека и гигиене окружающей среды РАМН и Минздравсоцразвития Российской Федерации / под ред. Ю.А. Рахманина, М., 2007). 1.2 Общие аспекты взаимодействия высокодисперсных веществ окружающей среды и растений Формирование современного облика нашей планеты из первозданного раскаленного шара в небесное тело с развитой биосферой во многом стало возможным благодаря деятельности живых организмов, которые трансформировали горные породы, создав почву. Это, отчасти, стало возможным 18 благодаря способности микроорганизмов и растений добывать питание из материнской породы разной дисперсности. Фактически весь период существования, так или иначе, организмы контактировали с микро- и наночастицами минерального происхождения. И именно способность растений к добыванию элементов питания из породы и ее фракций стало важным эволюционным признаком (Чижикова Н.П., Карпова Д.В. Минералы разных гранулометрических фракций как источники элементов питания растений (на примере агросерных тяжелосуглинистых почв Владимирского оползя) // Бюллетень Почвенного Института им. В.В.Докучаева. 2009. №63. С.41-49). В ходе эволюции растения непрерывно контактировали с высокодисперстными фракциями через воздух и адаптировались к такому воздействию. Так, по расчетам В.В Добровольского (Добровольский В.В. Химия жизни – М. Наука; 1988 – 176 с.) с каждого квадратного километра засушливых районов земли в атмосферу поступает примерно 100 тонн пыли, что, в совокупности, составляет около 6 млрд. тонн пыли ежегодно, захватываемой ветрами в атмосферу. Среднестатистическое содержание минеральных частиц различных размеров в атмосфере земли составляет более 20 млн. тонн, а в 1 м3 самого чистого атмосферного воздуха концентрируется не менее 1 млн. минеральных взвешенных частиц от 0,001 до 1000 мкм (Yushkin N.P. Mineralis mundi et biosphere: mineralis organiz-mobioz, biomineral interaction coevolution // acta iv International seminar “Mineralogy et vitam: Origin de biosphere et co-evolution of mineralis et biologicum mundos biomineral-ogiya” (Syrtyvkar, Komi Republie, May 22-25, 2007). Syrtyvkar. 2007. p. 5-7). Куда более активен контакт растений с высокодисперсными частицами через воду, в одном кубическом дециметре воды содержится 5 – 6 млн. таких частиц (Lisitsyn Q.P. Oceanic processibus sedimentation. Lithology et geochemistry. M.: Nauka, 1978. Р. 392; Kravchyshyn M.D. Materiales composition agueum suspension Northern Dvina aestuario (albus mare) dum vernum aestus // Oceanology. 2010. T.50. №5. Р. 396-416). Элементный состав частиц разнообразен. В 19 частности, океанская пыль представлена, преимущественно, гиалитом и сульфатами, континентальная – кварцем, сульфатами, углеродом, силикатами, алюмосиликатами и др. (Bogaticov O.A. Anorganicis nanoparticles in natura // Bulletin de RAS. 2003. T.73. №5. Р. 426-428; Amorim R. Zeoliti structures loading with an anticancer compound as drug delivery systems // Vournal of physical chemistry. 2012. V.116. Р. 25642 – 25650; Голохваст К.С., Никифоров П.А., Кику П.В., Чайка В.В., Автомонов Е.Г., Чернышев В.В. и др. Атмосферные взвеси Владивостока: Гранулометрический и вещественный анализ // Экология человека. 2013. №1. С. 14-19). Наиболее выражен контакт растений с высокодисперсными веществами через почву. В почве, в норме, содержится большое количество дисперсных частиц размером менее 1 мкм, представленные глинистыми минералами и остатками органических соединений. На долю тонкодисперсных частиц в почве приходится 10-20% массы, иногда до 50-60%. Это обстоятельство определяет уникальные сорбционные свойства почв, получившие обозначение как обменная поглотительная способность. Тонкодисперсные частицы притягивают своей поверхностью катионы, находящиеся в растворах. При этом между тонкодисперсными веществами и почвенными водами существует подвижное равновесие. Обменная поглотительная способность играет важную роль в обеспечении непрерывного перехода элементов в клетки растений. Этот процесс реализуется через вытеснение ионами водорода катионов находящихся в сорбированном состоянии на поверхности почвенных частиц. Путь катиона из кристаллохимической структуры минерала в растение может быть описан схемой: «минерал – раствор – поглощенное состояние – раствор – растение» (Добровольский В.В. Химия жизни. М., 1988. 176 с.). Вышеописанные события «в норме» разворачиваются при поступлении элементов минерального питания из почв. Между тем, окружающая среда растений, во многом благодаря деятельности человека насыщена, сегодня, и частицами искусственного происхождения. Основной вклад в содержание взвешенных в атмосфере техногенных частиц вносит транспорт, прежде всего, 20 автомобильный - в виде частиц шин дорожного полотна, сажи, и т.д. Значительная часть взвешенных в воздухе частиц имеет промышленное происхождение, особенно стройиндустрии, в том числе частицы цемента, извести, песка и т.д. или теплоэнергетики (Голохваст К.С., Христофорова Н.К., Кику П.Ф., Гульков А.Н. Гранулометрический и минералогический анализ взвешенных частиц в атмосферном воздухе // Физиология и потология дыхания. 2011. Вып.40. С.94-100). Взвешенные вещества частично могут поступать в атмосферу вследствие почвенной эрозии. Размерность взвесей значительно варьирует от 100 нм до 100 мкм. Преобладание того или иного класса частиц во многом определяется их происхождением. В США, начиная с 1990 года выявлен факт преобладания взвешенных в атмосферном воздухе частиц (57%) величиной менее 10 мкм (Repozt on a WHO working group meeting, 2004). В ходе исследований К.С. Головаст и др. (Голохваст К.С., Чекрыжов И.Ф., Паничев А.М., Кику П.В. и др. Первые данные по вещественному составу атмосферных взвесей Владивостока // Известия Самарского науного центра РАН. 2011. Т.13. №1(8). С.1853-1857) установлено преобладание в воздухе российских городов (Владивосток, Хабаровск и др.) частиц третьего размерного класса от 10 до 50 мкм (35 - 50%). В то же время в зоне вокруг ТЭЦ и ряда промышленных объектов авторы констатировали наличие многих фракций первого и второго размерного класса, от 0,1 до 1 мкм, и от 1 до 10 мкм. Значительный вклад в техногенное загрязнение окружающей среды вносит горнодобывающая и металлургическая отрасли. Масштабы этой деятельности таковы, что ряд исследователей предложили последние сто лет рассматривать как период металлизации биосферы. Безусловно, что это отразилось на растениях. В норме в атмосферном воздухе присутствует некоторое количество рассеянных металлов, их концентрация примерно в 100 раз меньше, чем в природных водах, но в совокупности их общая масса составляет несколько тысяч тонн для цинка, меди, сотни тонн для марганца, свинца и т.д. Пути техногенного рассеивания металлов многообразны. Значительны выбросы отходов в атмосферу при металлургических процессах. Так, например, 21 доменные газы уносят в атмосферу не только железо, но и медь, свинец, мышьяк и т.д. В пыли, выбрасываемой при плавке медных руд, содержится до 10% меди. Перед выплавкой металлов из руды, последняя проходит длительную подготовку, в ходе которой значительная часть металла уходит в пыль. Ультрадисперсные частицы разносятся ветром, попадают в природные воды и, в конечном итоге, входят в контакт с микроорганизмами и растениями. Масштабы техногенной миграции металлов настолько значительны, что только за 1965 – 1975 годы их величина превысила 600 тыс. тонн для меди, 500 тыс. тонн цинка, 300 тыс. тонн свинца и т.д. Вышесказанное демонстрирует важность всестороннего изучения роста, развития и адаптации растений к относительно «новым» техногенным компонентам окружающей среды. 1.3 Приспособление растений к воздействию техногенных наноматериалов. Взаимодействие в системе «наночастицы – растения» Наукой приспособления накоплен растений значительный к условиям багаж знаний среды обитания об с особенностях содержанием техногенных наноматериалов. Первые работы по данной тематике приходятся на 19 век и связаны с исследованиями действия сажи на растения в Великобритании по мере развития промышленного производства. В последующем получили развитие исследования по оценке действия цементной, асбестовой пыли на растениях. Накопленный материал указывает, что цементная пыль при попадании в почву замедляет рост большинства культурных растений (лук, чеснок, пшеница и др.). По мнению К.А. Пассика (Пассика К.А. Исследование влияния выбрасов цементной пыли на рост и развитие растений // Успехи современного естествознания. 2004. №11. 45 с.) это вызвано недостаточной аэрацией почвы изза образования пленки цементной пыли и увеличения ph почвы. Техногенное изменение эдафических условий и, соответственно, процессов минерального питания растений рассматривается в качестве одной из ведущих причин деградации лесов в окрестностях промышленных предприятий (Лукина 22 Н.В., Никонов В.В. // Лесоведение. 1999. №2. С.57-67: Helmissaar H.S., Makkonen K., Olsson M. et al Plant and soil. 1999. Р.193-200). Исходя из работы Г.А. Зайцева, Г.М. Мухаметовой, Д.В. Веселкина (Зайцев Г.А., Мухаметова Г.М., Веселкин Д.В. Особенности формирования микоризы сосны обыкновенной в условиях промышленного загрязнения // Вестник Оренбургского гос. ун-та. 2009. №6. С.137-139), одним из следствий этого является ослабление и полное подавление микоризообразования. Микориза – симбиотная ассоциация мицелия гриба с корнями высших растений. Детальное изучение техногенного давления на эктомикоризу проведено Д.В. Веселкиным (Веселкин Д.В. Реакция экомикориз Pinus sylvestris L. на техногенное загрязнение различных типов // Сибирский экологический журнал. 2005. №4. С.753-761; Веселкин Д.В. Соврменная микология в России // Первый съезд микология в России: тез. докл. М., 2002. С.86-87). Установлено увеличение объема эктомикоризных окончаний на загрязненных металлами территориях. Учитывая, что утолщение – один из известных симптомов токсического действия ионов меди на корни растений, возрастание поперечного размера корня в составе микориз может и не иметь адаптивного значения, являясь результатом токсического повреждения растительных тканей (Веселкин Д.В. Реакция экомикориз Pinus sylvestris L. на техногенное загрязнение различных типов // Сибирский экологический журнал. 2005. №4. С.753-761). Вполне очевидно, что уникальные свойства наноматериалов – проникающая способность через биологические мембраны, значительная удельная поверхность, а, соответственно, и многократно повышенная реакционная способность (Глущенко Н.Н. Физико-химические закономерности биологического действия высокодисперсных порошков металлов: автореф. дис. д-ра биол. наук, М.,1989; Рахметова А.А., Алексеева Т.П., Богословская О.А., Лейпунский И.О., Ольховская И.П., Жигач А.Н., Глущенко Н.Н. Ранозаживляющие свойства наночастиц меди в зависимости от их физико-химических характеристик // Российские нанотехнологии. 2010. Т.5. №3-4. С.102) уже в ближайшем будущем обеспечат доминирование наноформ на рынке микроудобрений и компонентов 23 для растениеводства. исследования по Подтверждением оценке этого биологических являются свойств многочисленные наноматериалов на биологических моделях (Арсеньева И.П. Аттестация наночастиц металлов, используемых в качестве биологически активных препаратов // Нанотехника. 2007. №10. С.72; Богословская О.А., Сизова Е.А., Полякова В.С., Мирошников С.А., Глущенко Н.Н. Изучение безопасности введения наночастиц меди с различными физико-химическими характеристиками в организм животных // Вестник Оренбургского государственного университета. Оренбург, 2009. №2. С. 124-127). В частности, для наночастиц металлов показана относительно низкая биотоксичность в сравнении с минеральными формами. Так, по данным Н.Н. Глущенко и др. (Глущенко Н.Н., Богославская О.А., Ольховская И.П. Сравнительная токсичность солей и наночастиц металлов и особенность их биологического действия // материалы междунар. науч.-практ. конф. Нанотехнологии и информационные технологии – технологии 21 века. М.: Наука, 2006. С. 93-95) LD 50 сульфата меди в 7,5 раз выше в сравнении с наночастицами меди (d = 100 нм). Аналогичная разница для цинка составляет 28 раз, железа - 36 раз. Данные факты послужили основанием для использования наночастиц металлов в практике. Л.В. Коваленко, Г.Э. Фолманис (Коваленко Л.В., Фолманис Г.Э. Биологически активные нанопорошки железа. М.: Наука, 2006. 124 с.) установлено, что предпосевная обработка семян нанопорошками железа в концентрации 0,001 % положительно влияла на энергию прорастания, однако увеличение концентрации до 0,01 % приводило к подавлению прорастания. Была рассчитана оптимальная доза предпосевной обработки, дающей от 5 до 30 % повышения урожайности и улучшения товарного вида растительной продукции. Широкое распространение наночастицы металлов получили в растениеводстве при предпосевной обработке семян. Одним из первых кто предложил этот метод был С.В. Крылов, который заменил ядохимикаты на наночастицы при протравливании, в частности, пестицида фентиурама. На основании проведенных исследований и последующих испытаний им был создан препарат, включающий 24 гидрофобный полимер наночастиц металлов, которые защищают семена и всходы от патогенов и стимулируют рост и развитие растений, сокращающие сроки вегетации (Крылов С.В.). Автором в эксперименте были выявлены оптимальные соотношения компонентов, в т.ч. наночастиц металлов (Cu, Fe, Zn, Mo), описаны обеззараживающие и стимулирующее действие наноматериалов при обработке семян (Крылов С.В., Паничкин Л.А., Захарин А.А. и др. Способ оценки метода протравливания семян // Патент РФ №2015631 от 15.07.1994 г; Черняев Н.Г. Изучение влияния препаратов и способов обработки семян на перезимовку озимых зерновых культур // Известие ТСХА. 1997. Вып.2. С. 23-28; Райкова А.П. Предпосевная обработка семян // материалы междунар. науч.-практ. конф. «Льняной комплекс россиии, проблемы и перспективы. Вологда, 2001; Райкова А.П., Паничкин Л.А., Райкова Н.Н. Использование ультрадисперсных порошков металлов для предпосевной обработки семян // Доклады ТСХА. 2004. Вып. 276. С. 44-48; Райкова А.П., Паничкин Л.А., Райкова Н.Н. Исследования влияния ультрадисперсных порошков металлов, полученные различными способами на рост и развитие растений // материалы междунар. науч.-техн. конф. «Нанотехнологии и информационные технологии – технологии 21 века». М., 2006. С. 118-123). Л.А. Паничкиным и А.П. Райковой (Паничкин Л.А., Райкова А.П. Использование нанопорошков металлов для предпосевной обработки семян сельскохозяйственных культур // Известия ТСХА. 2009. Вып.1. С. 59-65) показано благоприятное действие предпосевной обработки семян пшеницы, ячменя и т.д. нанопорошками Cu, Fe, Zn на урожайность и содержание белка. Авторами рекомендовано использовать данные вещества для опрыскивания вегетирующих растений и прикорневой обработки с целью защиты от гельминтоспориоза, пероноспороза, аскохитиза и др. Предпосевная обработка семян наночастицами железа, кобальта и меди оказывает положительное действие на формирование ассимилирующего аппарата растений рапса и кукурузы (Сушилина М.М. Влияние ультрадисперстных порошков металлов (УДПМ) – новых микроудобрений на урожайность и качество зеленой массы: автореф. дис. канд. биол. наук: 25 06.01.2004. М., 2004. 25 с.; Сушилина М.М., Монькина А.И. Нанотехнологии в растениеводстве и сельском хозяйстве // Вестник ФГБОУ ВПО РГАТУ. 2011. №3(11). С.42-44). Так, в производственных опытах изучено влияние предпосевной обработки семян кукурузы наночастицами железа на интенсификацию роста и развития, повышение урожайности зеленой массы и зерна этой культуры. В результате происходила интенсификация фотосинтетических процессов. Содержание Fe, Cu, Mn, Cd и Pb в онтогенезе кукурузы варьировало в широких пределах, но адсорбция наночастиц Fe на начальных стадиях развития растений влияла на уменьшение содержания этих химических элементов в созревающем зерне, чему сопутствовало Физиологически действие изменение отдельных его биохимических наночастиц на семена свойств. растений определяется их влиянием на активность фитогормонов. Показано, что предпосевная обработка семян вики и пшеницы наночастицами меди до 10 т/га сопровождается увеличением содержания цитокина и индолилуксусной кислоты. Практика использования наночастиц может носить и достаточно специфический характер. Так, оценка эффективности препаратов наночастиц серебра и тиосульфата серебра, при сохранении срезанных гвоздик (Dianthus caryophyllus), показала, что курс «лечения» наночастицами серебра и тиосульфатом серебра продлевает долговечность срезанных гвоздик за счет уменьшения окислительного стресса (Врублевский Э., Киреев В., Недзвецкий В., Сосновцев В. Нанотехнология – путь в будущее или бренд для финансирования // Нано- и микросистемная техника. 2007. № 12. С. 6–20). Наноматериалы получают свое применение при производстве обогащенного мироэлементами растительного сырья. Это стало возможным благодаря аккумуляции наночастиц растениями. В исследованиях на растениях риса показано увеличение содержания наночастицы меди в корнях, коре и листьях растений. Разработан новый способ обогащения селеном растительного сырья с применением водных суспензий не взаимодействующих с водой элементов, в первую очередь, селена. Исследовано влияние водной суспензии наноразмерного 26 селена на подсолнечник румынского сорта «Малыш» (Bogaticov O.A. Anorganicis nanoparticles in natura // Bulletin de RAS. 2003. T.73. №5. Р. 426-428). Показано, что обработка наземной части топинамбура во время вегетационного периода препаратами, содержащими селен как в виде наночастиц, так и в виде солей селена, повышает содержание селена во всех частях растения. Установлено, что селен преимущественно концентрируется в листьях растения. Уникальные свойства наночастиц предопределили широкое применение последних для трофических целей помимо растениеводства в различных областях животноводства (Куренева Е.Н., Егоров И.А., Федоров Ю.И., Глущенко Н.Н., Фаткулина Л.Д. Использование высокопродуктивных металлов в составе премиксов комбикормов для бройлеров // Новое в кормлении и содержании сельскохозяйственной птицы. Загорск, 1984. С. 3-8; Егоров И.А., Куренева В.П., Глущенко Н.Н., Фаткуллина Л.Д., Федоров Ю.И. Высокодисперсные порошки металлов – источники микроэлементов для сельскохозяйственной птицы // Физиолого – биохимические основы повышения продуктивности сельскохозяйственной птицы // Сборник научных трудов. Т.31. Боровск, 1985. С.80-88; Ильичев Е., Назарова А., Полищук С., Иноземцев В. Переваримость рациона и баланс питательных веществ при скармливании телятам нанопорошков кобальта и меди // Молочное мясное скотоводство. 2011. №5. C. 27-29; Мирошникова Е.П., Аринжанов А.Е., Глущенко Н.Н., Василевская С.П. Обмен химических элементов в организме карпа при использовании наночастиц кобальта и железа в корме // Вестник Оренбургского государственного университета, 2012. №6. С. 55-57; Мирошникова Е.П., Аринжанов А.Е., Килякова Ю.В. Изменение гематологических параметров карпа под влиянием наночастиц металлов // Достижения науки и техники АПК. 2013. №5. С. 55-57; Аринжанов А.Е., Мирошникова Е.П., Килякова Ю.В. Воздействие наночастиц комплекса металлов на организм карпа // Известия Оренбургского государственного аграрного университета. 2013. №2. С. 113-116). Между тем, одной из сдерживающих причин дальнейшего активного использования наночастиц в сельском хозяйстве стали пионерские работы по нанотоксикологии. «Пилотные» исследования по данной 27 проблематике выявили факты выраженного токсического действия отдельных наноформ (Chen Z. Acute toxicological effects of copper nanoparticles in vivo // Toxicology Letters 2006. Vol. 163. Iss. 2. P. 109-120). Так, на модели теплокровных животных V.E. Kagan et al (2006) показано, что с попаданием в легкие углеродных нанотрубок размером меньше 10 нм постепенно развиваются злокачественные новообразования. Нельзя сказать, что человечество впервые столкнулось с пагубными действиями наноматериалов на живое. Еще в древние времена люди, занятые выплавкой цинка, страдали от цинковой лихорадки, вызванной вдыханием аэрозолей окиси цинка. Уже в 18-19 веках было известно пагубное действие асбестовой пыли, высокодисперсной сажи на биосферу. Существуют данные о снижении адаптационных свойств растений на фоне дополнительного поступления наночастиц извне. Опираясь на накопленные к настоящему моменту знания о свойствах различных наночастиц, можно выделить основные причины их токсического воздействия на живые системы, в общем, и растения, в частности. Во-первых, токсичным может быть основное вещество наночастицы, как, например, в случае «квантовых точек» – именно Cd2+, токсичность которого велика как в нано-, так и в макросостоянии, является причиной токсичности для растительных клеток. Вовторых, даже если наночастицы сами по себе биологически безвредны, некоторые из них могут выступать в роли катализаторов образования токсичных веществ, катализируя фотоокисление, вирусы. Специфическое воздействие вещества на организм может быть обусловлено непосредственно тем, что оно находится в наносостоянии. фторопласт, Например, широко химически используемый инертный для и безопасный изготовления посуды, полимер будучи распыленным в воздухе в виде наночастиц диаметром 26 нм в ничтожной концентрации (60 мкг/м3) способен убить крысу за 30 минут, вызывая кровоизлияния в легких. Механизмы развития токсического эффекта наночастиц разнообразны (Reevesa J.F., Daviesa S.J., Dodda N.J.F., Jha A.N. Hydroxyl radicals (OH) are associated with titanium dioxide (TiO2) nanoparticle-induced cytotoxicity and oxidative DNA damage in fish cells // Mutation Research/Fundamental and Molecular 28 Mechanisms of Mutagenesis 2008. Vol. 640. Iss. 1-2. P. 113-122; Kaura I.P., Bhanarib R., Bhanarib S., Kakkara V. Potential of solid lipid nanoparticles in brain targeting // J. of Controlled Release 2008. Vol. 127. Iss. 2. P. 97-109). Наночастицы способны проходить сквозь обычные защитые барьеры живых организмов и инициировать различные клеточные реакции: пролиферацию клеток и нарушение клеточного цикла, апоптоз (Unfried K., Sydlik U., Bierhals K., Weissenberg A., Abel J. // Am.J. Physiol. Lung Cell Mol. Physiol, 2008. V. 294. № 2. P. L358; Xia T., Kovochich M., Liong M., Zink J.I., Nel A.E. // ACS Nano. 2008. V. 2. № 1. P. 85; Atha D.H., Wang H., Petersen E.J., Cleveland D., Holbrook R.D., Jaruga P., Dizdaroglu M., Xing B., Nelson B.C. Copper oxide nanoparticle mediated DNA damage in terrestrial plant models // Environ. Sci. Technol. 2012. Vol. 46, P. 1819−1827; Park E.J., Kim H., Kim Y., Park K. // Arch. Pharm. Res. 2010. V. 33. № 5. P. 727). Невозможность адаптации растений к воздействию наночастиц, равно как и снижение способности к приспособлению, обусловлена повреждением мембраны клетки и различных клеточных структур, нарушением структуры белков и изменение их функциональной активности, повреждением ДНК, изменением экспрессии генов (Meng H., Xia T., George S., Nel A.E. // ACS Nano. 2009. V. 3. № 7. P. 1620; Nel A.E., Madler L., Velegol D., Xia T., Hoek E.M., Somasundaran P., Klaessig F., Castranova V., Thompson M. Understanding biophysicochemical interactions at the nano-bio interface // Nat. Mater. 2009. Vol.8. P. 543—557; Подколодная О.А., Игнатьева Е.В., Подколодный Н.Л., Колчанов Н.А. Пути поступления наночастиц в организм млекопитающих, их биосовместимость и клеточные эффекты // Успехи современной биологии. 2012. №1. С. 3-15). Между тем, рассматривая накопленный экспериментальный материал, можно обнаружить, что не всегда и не везде наноматериалы оказывают токсическое или иное повреждающее действие. Так, одни исследователи однозначно обнаружили цитотоксический эффект магнитных частиц на основе оксида железа, другие же напротив, показали, что они безвредны (Prow T., Smith J.N., Grebe R., Salazar J.H., Wang N., Kotov N., Lutty J., Leary J. Construction, gene 29 delivery, and expression of DNA tethered nanoparticles // Molecular Vision 2006. Vol. 12. P. 606-615). Особенности действия наночастиц на живые организмы определяются условиями среды. Так, исследования токсичности наночастиц кадмия, хрома, меди, никеля и цинка на водной культуре дафний (Daphnia magna) показали, что проявления токсических свойств этих веществ зависит от значений рН среды (Van Sprang P.A., Janssen C.R. Toxicity identification of metals: development of toxicity identification fingerprints // Environmental Toxicology and Chemistry. 2001. Vol. 20. Iss. 11. P. 2604–2610). Воздействие различных концентраций суспензий микрочастиц, наночастиц и ионов цинка на водные культуры дафний (Daphnia magna) и бактерий (Vibrio fischeri) выявило летальные концентрации (ЛК50) – 8,8, 3,2 и 6,1 мг/л для дафний и 1,8, 1,9 и 1,1 мг/л для бактерий, соответственно. Сильными токсическими свойствами обладают наночастицы алюминия, которые способны подавлять синтез РНК и вызывать нарушение функций митохондрий в клетках и т.д. В тоже время одним из основных механизмов токсического действия является окислительный стресс, который обуславливается активными формами кислорода, генерируемыми наночастицами. Несмотря на особое внимание к проблеме биологических эффектов наночастиц металлов, есть серьезный недостаток информации относительно их влияния на экосистемы. Накопленные наукой факты предопределили образование в 2008 году Международного нанотоксикологического альянса по гармонизации требований к охране здоровья и окружающей среды (International alliance for nanoENS Harmonization, IANH) в целях разработки протоколов тестирования наноматериалов на клеточном и организменном уровнях. Вопросы, связанные с этикой использования искусственных наноматериалов и регулирования данной деятельности активно разрабатываются подготовлена «Концепция и в идентификации и России. Роспотребнадзором количественное определение наноматериалов», утвержденная постановлением главного государственного санитарного врача РФ № 29 от 31.10.07 (Онищенко Г.Г. Организация надзора за оборотом наноматериалов, представляющих потенциальную опасность для 30 здоровья человека // Гигиена и санитария. 2011. №13. С. 164-173). Значительное внимание в «концепции» уделено оценке токсичности наноматериалов на растительных объектах. Рассматривая различные сценарии воздействия наночастиц на растения, следует детально изучить взаимодействие, последних, с биологическими объектами. Как следует из результатов исследований (A.E. Nel, L. Madler, D. Velegol et al (Nel A.E., Madler L., Velegol D., Xia T., Hoek E.M., Somasundaran P., Klaessig F., Castranova V., Thompson M. Understanding biophysicochemical interactions at the nano-bio interface // Nat. Mater. 2009. Vol.8. P.543—557), наночастицы могут встраиваться в мембраны, проникать в клетки и клеточные органеллы, взаимодействовать с нуклеиновыми кислотами, белками, что в результате, может изменять функции различных биологических структур. В этой связи, высокая биологическая активность наночастиц определяется возникновением их комплексов с белками (Maiorano G., Sabella S., Sorce B., Brunetti V., Malvindi M.A., Cingolani R., Pompa P.P. // ACS Nano.V. 4. № 12. P. 7481). В зависимости от аффинности белки короны либо «жестко» связаны с частицей, либо находятся в состоянии динамического обмена между средой и наночастицей. Время жизни комплекса может варьировать от миллисекунды до десятков часов (Nel A., Xia T., Madler L., Li N. // Science. 2006. V. 311. № 5761. P. 622; Ehrenberg M.S., Friedman A.E., Finkelstein J.N., Oberdorster G., McGrath J.L. // Biomaterials. 2009. V. 30. № 4. P. 603). Это, в конечном итоге значительно снижает адаптационные способности растений. Состав короны определяется локальными физико-химическими свойствами наноматериала (Lundqvist M., Stigler J., Elia G., Lynch I., Cedervall T., DawsonlK.A. // Proc. Natl. Acad. Sci. USA. 2008. V. 105. № 38. P. 14265). Макромолекулы, адсорбированные на поверхности наночастицы, изменяют ее физические свойства, также как поверхностный заряд, размер, стабильность, что отражается на механизмах поглощения наночастиц клетками, определяет их распределение в органах и интенсивность выведения из него (Linse S., Cabaleiro-Lago C., Xue W.F., et al. // Proc. Natl. Acad. Sci. USA. 2007. V. 104. № 21. P. 86-91; Cabaleiro-Lago C., 31 Quinlan-Pluck F., Lynch I., Lindman S., Minogue A.M., Thulin E., Walsh D.M., Dawson K.A., Linse S. // J. Am. Chem. Soc. 2008. V. 130. № 46. P. 154; Wu W.H., Sun X., Yu Y.P., Hu J., Zhao L., Liu Q., Zhao Y.F., Li Y.M. // Biochem. Biophys. Res. Commun. 2008. V. 373. № 2. P. 315). Изменение свойств белков при контакте с наночастицами можно проиллюстрировать данными S.S. Karajana и др. (Karajanagi S.S., VertegelA.A., Kane R.S., Dordick J.S. // Langmuir. 2004. V.20. №26. P.115), свидетельствующими о потери до 99% активности фермента альфа-химотрипсина после образования комплекса с однослойными однородными нанотрубками. При адсорбции белков на наночастицах большего размера наблюдаются более выраженные изменения структуры и функции белков (Vertegel A.A., Siegel R.W., Dordick J.S. // Langmuir. 2004. V. 20. № 16. P. 68). Останавливаясь на механизмах проникания наночастиц в растительные клетки, можно отметить их разнообразие - от эндоцитоза, с участием клатрина и кавеол, до диффузии и других механизмов запускаемые электростатическими силами, Ван-дер-Ваальсовыми, либо стерическими взаимодействиями (Raffa V., Ciofani G., Vittorio O., Riggio C., Cuschieri A. // Nanomedicine. 2010. V. 5. № 1. P. 89). Механизм проникновения наночастиц в клетку зависит от свойств самой частицы (заряд, размер, форма, способность формировать агломераты т.д.), особенностей клеток (наличие специализированных структур, обеспечивающих тот или иной тип захвата частиц), микроокружения (наличие тех или иных компонентов, способных модифицировать поверхность частицы). Интенсивность поглощения клетками заряженных наночастиц выше, чем у нейтральных. Причем, положительно заряженные частицы поглощаются более эффективно, чем заряженные отрицательно (Panyam J., Zhou W.Z., Prabha S., Sahoo S.K., Labhasetwar V.// FASEB J. 2002. № 16. P. 1217; Foged C., Brodin B., Frokjaer S., SundbladA. // Int. J. Pharm. 2005. V. 298. № 2. P. 315; He C., Hu Y., Yin L., Tang C., Yin C. // Biomaterials. 2010. V. 31. № 13. P. 36-57; Подколодная О.А., Игнатьева Е.В., Подколодный Н.Л., Колчанов Н.А. Пути поступления наночастиц в организм 32 млекопитающих, их биосовместимость и клеточные эффекты // Успехи современной биологии. 2012. 132. №1. С. 3-15). Учитывая, что наиболее активен контакт наноматериалов с высшими растениями через корневую систему, закономерно, что именно на этом этапе растения выработали механизмы защиты от негативного действия этих веществ. Это подтверждается исследованиями последних 5-7 лет. В исследованиях по аккумуляции и распределению наночастиц платины в органах водных и почвенных культур одно- и двудольных растений - пшеницы и гороха установлено, что 10-дневные водные культуры накапливали наночастицы платины больше, чем почвенные, а корни - значительно больше, чем стебель и листья. У пшеницы водной культуры в большей степени выражена аккумулирующая способность, чем у аналогичных растений гороха, а пшеница почвенной культуры, наоборот, с меньшей интенсивностью накапливала (поглощала) наночастицы платины, чем почвенная культура гороха. При этом развитие адаптационной реакции растений на действие техногенных материалов может сопровождаться частичной или полной потерей корневой системы. На различных видах растений при использовании хлорат калия (KAuCl4), опосредованным наночастицами золота (Au), выявлено, значительное снижение длины и падение активности меристемы корней. В исследованиях по оценке действия KACl4 в совокупности с железом на корни арабидопсиса (Arabidopsis thaliana) выявлен факт потери растениями корневой системы. В тоже время подбором наночастиц и варьированием их дозировок можно добиться и совершенно иной адаптационной реакции растений. Проиллюстрировать это можно результатами исследований Г.Р. Кудояровой и др. (Кудоярова Г.Р., Трапеников В.К., Иванов И.И. Гидравлическая проводимость корней при гетерогенном распределении элементов минерального питания // Известия Уфимского научного центра РАН. 2013. №2. С. 33-37), которые констатировали факт стимулирующего действия наночастиц концентрации 0,01-1,0 мг/дм3 на развитие корневой системы. серебра в 33 Между тем, размер наночастиц также оказывает влияние на поглощение их клетками: чем меньше частица, тем скорость поглощения ее выше (Fang C, Shi B, Pei Y.Y., Hong M.H., Wu J., Chen H.Z. // Eur. J. Pharm. Sci. 2006. V. 27. № 1. P. 27; Lohbach C., Neumann D., Lehr C.M., Lamprecht A. // J. Nanosci. Nanotechnol. 2006. V. 6. № 9-10. P. 33). В соответствие с этим, относительно большая удельная поверхность наночастиц определяет большую реакционную способность, протекающую на границе «наноматериал и среда», что в отдельных случаях выражается и большей токсичностью. Генерация свободных радикалов и оксидитивный стресс являются одними из основных механизмов токсичности наноматериалов (Nel A., Xia T., Madler L., Li N. // Science. 2006. V. 311. № 5761. P. 622). По О.А. Подколодной и др. (Подколодная О.А., Игнатьева Е.В., Подколодный Н.Л., Колчанов Н.А. Пути поступления наночастиц в организм млекопитающих, их биосовместимость и клеточные эффекты // Успехи современной биологии. 2012. №1. С. 3-15) в генерации свободных радикалов могут быть задействованы различные механизмы: формирование активных донорно-акцепторных групп, образование гидроксильных радикалов, генерация их под действием света. Однако, безусловно, что биологические эффекты наночастиц при воздействии на растение более разнообразны и не укладываются в рамки той или иной классификации. Следовательно, рассматривая подобные взаимодействия, необходимо детально изучать весь спектр возможных эффектов. Это в полной мере относится к любому исследованию по оценке адаптации растений к определенный условиям среды, содержащей интерес могут представлять наночастицы. исследования В этой по связи, сравнению наночастиц вещества с другими его формами – ионной, макроскопической и т.д., что, например, было сделано при оценке, влияния ионов меди и наночастиц оксидов меди на Elоdeadensa Planch. Показано, что наночастицы более активно поглощались растениями элодеи. Обе формы меди активировали процессы перекисного окисления липидов (наночастицы - на 180%, ионы - на 120%). Активность каталазы и супероксиддисмутазы была повышенной (в 1,5 – 2,0 раза) 34 в вариантах с наночастицами. Ионы меди подавляли фотосинтез при концентрации 0,5 мг/л, тогда как наночастицы - при 1,0 мг/л. Закономерно, что невозможность адаптации растений к действию отдельных техногенных нанометериалов предъявляет новые требования к практике их использования и защите окружающей среды. В этой связи актуальными представляются исследования на стыке наук ботаники и экологии по определению пределов толерантности и оценке устойчивости растений к воздействию наноматериалов. Это направление в науке в последние десять лет получило значительное развитие. Исследования (Raphanussativus), толерантности рапса кукурузы (Brassicanapusnapus), (Zeamays огурца L.), редиса (Cucumissativus) к возделыванию наночастиц цинка и его оксида показали, что их концентрация 2000 мг/л отрицательно действует на прорастание семян кукурузы и подавляет удлинение корней. Пятидесятипроцентная ингибиторная концентрация (IC50) для редьки составила 50 мг/л, рапса – 20 мг/л (Lin D. Phytotoxicity of nanoparticles: inhibition of seed germination and root growth // Environmental Pollutants 2007. Vol. 150. Iss. 2. P. 243-250). Описаны хорошо выраженные межвидовые различия в адаптационных реакциях растений и их толерантности к действию наноматериалов. Влияние водных суспензий наночастиц TiO2 (размером 5 нм) и Al2O3 (размером 7 нм), в различных концентрациях, на растения фасоли обыкновенной, пшеницы мягкой яровой показало неоднозначное воздействие наночастиц на рост и развитие изученных растений, а также содержание хлорофиллов a, b и каротиноидов в листьях. Установлено, что наиболее устойчивой культурой к действию данных наночастиц является фасоль. Растения пшеницы характеризуются выраженной избирательной чувствительностью к исследованным наночастицам, что определяет специфику изменений физиологических показателей, в том числе повышение урожайности у растений после воздействия суспензией наночастиц TiO2. 35 Фитотоксичность наночастиц карбида кремния по отношению к растениям различна и наиболее выражена для овса, ячменя и пшеницы (Голохваст К.С., Памирский И.Э., Бородин Е.Н., Рева Г.В. и др. Влияние микрочастиц минералов на работу физиологических и биологических систем // Фундаментальные исследования. 2013. №6. С. 909-912). Между тем адаптация растений к действию различных наночастиц одного и того же элемента, равно как и различных его форм протекает различно. Это хорошо видно из результатов экспериментов с использованием сферических наночастиц железа FeО (d= 80 ± 5 нм), магнетита Fe3O4 (d=50-80 нм) и раствора сульфата железа (II) FeSO4 на всхожесть, длину листьев и содержание фотосинтетических пигментов хлорофилла, а, хлорофилла b и каротиноидов у проростков Triticum vulgare Vill. Наибольшая чувствительность проростков регистрировалась при увеличении концентрации сульфата железа (II) в среде, что проявлялось в снижении всхожести, ингибировании роста листьев и содержания ФП (Mtiller L., Riediker M., Wick P., Mohr M. // J. R. Soc. Interface. 2010. V. 7. P. 27). Адаптационные способности растений к воздействию наноформ железа неоднозначны для каждого изученного показателя: по показателю длины листьев проростков, растения наиболее восприимчивы к наночастицам железа; по содержанию общей суммы фотосинтетических пигментов токсичны наночастицы магнетита и железа. Максимальное содержание фотосинтетических пигментов зафиксировано при концентрациях 0,125 и 1*10-3 г/л наночастиц Fe0, 0,5 и 1*106 г/л наночастиц Fe3О4 и 1*10-3 г/л FeSO4. В эксперименте на модели салата-латук (Lactuca sativa) и люцерны (Medicago sativa) при культивировании в среде, содержащей наночастицы меди, оксид и гидроксид меди показано, что приспособление растений к наноформам вещества протекает наименее успешно. Вследствие воздействия наночастиц, длина корня снижалась у обоих растений. Кроме того, наблюдалось снижение содержание питательных веществ и активности ферментов. Имело место снижение активности каталазы в люцерне (корень и побег) и повышение 36 активности аскорбатпероксидазы в корнях обоих видов растений (Hong S., Bielinska A.U., Mecke A., Keszler B., Beals J.L., Shi X., Balogh L., Orr B.J., Baker J.B., Banaszak M.M. Interaction of Poly(amidoamine) Dendrimers with Supported Lipid Bilayers and Cells: Hole Formation and the Relation to Transport // Bioconjugate Chemistry 2004. Vol. 15. Iss. 4. P. 774–782). Механизмы проникновения, распределения, накопления и деградации в растительных организмах, а так же их взаимодействия с биологическими объектами (ДНК, белки и др.) в значительной степени определяется свойствами наночастиц. В этой связи, крайне актуальными представляются научные исследования, направленные на изучение специфических особенностей адаптации растений к присутствию в окружающей среде техногенных наночастиц металлов. Накопленные знания позволят разработать новые методы оценки токсического действия наноматериалов и использовать биологические эффекты наночастиц, обеспечивающие ростостимулирование растений. 1.4. Резюме Анализ научной литературы позволяет констатировать, что высшие растения на протяжении всей своей истории активно взаимодействовали с ультрадисперсными чужеродными частицами. Данное обстоятельство обусловило формирование у них специфических механизмов адаптации, которые вполне эффективны при приспособлении к действию минеральных частиц. Однако эти барьеры не являются преградой для техногенных наноматериалов, проникновение которых в растительную клетку осуществляется, в том числе при образовании комплексов частиц с белками и другими органическими соединениями. В результате наночастицы могут накапливаться и длительное время сохраняться в растениях, что снижает их адаптационные способности. Перечень только зарегистрированных наноматериалов превысил несколько тысяч наименований, их поступление в биосферу закономерно возрастает. Исследования последних 7-10 лет демонстрируют многогранность действия 37 наночастиц различного происхождения на растения, описаны адаптационные изменения в растительных организмах на фоне действия наноматериалов. При этом наряду с данными о снижении адаптационных способностей растений к действию наноматериалов, в литературе представлены материалы о стимулирующем действии наноформ вещества, однако, накопленные наукой данные далеко не полные и нередко противоречивы. В этой связи, для ботаники как науки, «изучающей строение растительных организмов, их рост и развитие, приспособление к условиям окружающей среды», важным и перспективным является детальное изучение адаптации растений к воздействию наночастиц металлов. Значительный интерес представляет также определение пределов толерантности и оценки устойчивости растений к наночастицам различного элементного соства, в том числе наночастицам металлов. 38 ГЛАВА 2. МАТЕРИАЛЫ И МЕТОДЫ ИССЛЕДОВАНИЯ 2.1 Объекты исследования 2.1.1. Растительные тест – объекты Исследования строения, роста и развития растений, их толерантности и адаптации к среде обитания, содержащей наночастицы металлов – микроэлементов (Cu, Fe) выполнены в период 2009-2014 годов на базе лаборатории «Агроэкологии техногенных наноматериалов» Всероссийского НИИ мясного скотоводства. Основаным объектом исследования являлись семена яровой мягкой пшеницы Triticum aestivum L. (сорт «Учитель»), соответствующие 1 классу, необработанные документами. протравителями Отбор обеспечивающим семян и удостоверенные производился формирование по соответствующими стандартным репрезентативных групп методикам, сравнения из идентичного по происхождению, массе, всхожести и другим, предварительно оцененным признакам, экспериментального материала. Предназначенные для проращивания семена хранили в защищенном от света и влаги месте; пред проведением экспериментов их предварительно прогревали при температуре 3040°С в течение 5-7 суток в термостате. В ряде экспериментов объектом воздействия наночастиц меди и железа являлись луковицы Allium cepa L. (сорт «Штутгартен») массой 10-20 г и диаметром 1,5-2 см. 2.1.2. Бактериальные тест – объекты С целью уточнения и детализации биологических эффектов наночастиц и других соединений Cu и Fe были выполнены дополнительные эксперименты с использованием в качестве тест-объектов генноинженерных люминесцирующих микроорганизмов. 39 Первым подобным объектом был сенсорный штамм Escherichia coli K12 TG1, конститутивно экспрессирующий luxCDABE-гены морской бактерии Photobacterium leiognathi 54D10 и выпускаемым под коммерческим названием «Эколюм» (НВО «Иммунотех», Россия). В результате подобной генетической организации ферментная система генерации свечения (биолюминесценции) оказывалась тесно интегрированной с основными энергетическими потоками бактериальной клетки, вследствие чего токсическое воздействие закономерно вело к снижению интенсивности бактериального свечения. Второй группой микробных объектов являлись репортерные штаммы на основе E.coli K12 MG1655, несущие плазмиды с генными слияниями soxS::lux, katG::lux, recA::lux и любезно предоставленные д.б.н. И.В.Мануховым (ГосНИИгенетика, Россия). В результате подобной генетической организации названные штаммы характеризовались относительно низким уровнем свечения и специфично отвечали его индукцией при воздействии супероксид-аниона, перекиси водорода или повреждении ДНК несущих их клеток. 2.1.3. Наночастицы и другие соединения меди и железа 1 Наночастицы (рисунок 1), полученные в Институте энергетических проблем химической физики РАН (Москва) на установке МиГен методом высокотемпературной конденсации: 1.1 Наночастицы меди - сферические частицы (диаметр 103±2 нм), покрытые тонкой оксидной пленкой из CuО. Состав: медь кристаллическая - 96%, меди оксид – 4 %. 1.2 Наночастицы железа - сферические частицы (диаметр 80±15 нм), покрытые толстой оксидной пленкой из Fe3O4, α -Fe2O3 и γ- Fe2O3. Состав: железо кристаллическое – 85 %, оксиды железа – 15 %. 2 Микрочастицы. 2.2 Микрочастицы меди – сферические частицы (размер 40 микрон, чистота 99,5%). Производитель: Sigma – ALDRICH Chemie Gmbh, Riedstr (Германия); cat. No: 26,608-6. 40 Рисунок 1 – Типичные электронно-микроскопические изображения наночастиц железа и меди, полученные методом ПЭМ (слева) и СЭМ (справа). 2.1 Микрочастицы железа – сферические частицы (размер < 10 микрон, чистота 99,5%). Производитель: Alfa Aesar Gmbh (Германия); cat. No: 231-096-4. 3 Ионные формы Cu и Fe (соли): 3.1 Медь сернокислая 5-водн. (CuSO4 × 5H2O), ч.д.а.., ГОСТ 4165-78. Производитель: ЛенРеактив; Россия, Санкт-Петербург. 3.2 Железо сернокислое 7-водн., (FeSO4 × 7H2O), ч.д.а., ГОСТ 4148-78. Производитель: ЛенРеактив; Россия, Санкт-Петербург. Перед исследованием влияния нано- и микрочастиц или солей Cu и Fe их навески в количестве, соответствующем концентрациям 100 ммоль/л, помещали в стеклянные емкости, куда вносили по 10 мл химически чистой дистиллированной воды, после чего диспергировали путем обработки ультразвуком частотой 35 кГц в источнике ванного типа «Сапфир ТТЦ» (ЗАО ПКФ «Сапфир», Россия) в течение 30 мин. На основе полученных суспензий или растворов готовили 4-кратные разведения с концентрациями от 100 до 0,39 ммоль/л в пересчете на Cu или Fe. 41 2.2 Методы исследования 2.2.1 Методы исследования с использованием растительных организмов 2.2.1.1 Культивирование растительных организмов в контакте с наночастицами меди и железа для оценки их толерантности к подобному воздействию Планирование и выполнение исследований соответствии с Методическими указаниями МУ были осуществлены в 1.2. 2635-10 «Медико- биологическая оценка безопасности наноматериалов». – М.: Федеральный центр гигиены и эпидемиологии Роспотребнадзора, 2010. Суть исследваний заключалась в оценке толерантности растений к присуттвию в среде культивирования наночастиц и сравниваемых соединений меди и железа. Оборудование, материалы, реактивы: - Весы лабораторные общего назначения ГОСТ 24104-2001. - Анализатор жидкости «Эксперт–001». - Термостат ТС-1/80 СПУ ТУ 9452-02-00141798-97. - Ультразвуковой дезинтегратор. - Лупа «Микромед» МС-1. - Холодильник бытовой, обеспечивающий хранение проб (от +2 до +4 °С), ГОСТ 26678-85. - Пипетки автоматические дозаторы (любого типа), емкостью 1,0 см3 , ТУ 9452-002-33189998-2002. - Колбы мерные ГОСТ 1770-74. - Вода дистиллированная ГОСТ 6709-72. В качестве субстрата для выращивания Triticum aestivum использовали кварцевый песок, который промывали и просеивали для получения фракции 0,52,0 мм. С целью удаления примесных элементов песок замачивали в 10% растворе 42 HCl в течение 24 ч, после чего 20-кратно промывали дистиллированной водой до достижения нейтрального значения pH, контролируемого с использованием анализатора жидкости «Эксперт–001» (ООО «Эконикс-эксперт», Россия). Песок повторно высушивали и просеивали, после чего прокаливали в сушильном шкафу при температуре 130º С в течение 1 часа. Навески песка помещали в герметичные пластиковые контейнеры, после чего увлажняли лиозолями наночастиц или суспензиями микрочастиц, растворами солей Cu и Fe до полной влагоемкости, определенной по ГОСТ 12038-84. В качестве контролей использовали образцы песка, увлажненные тем же количеством дистиллированной воды. Дизайн эксперимента и использованные дозировки тестируемых соединений представлены в таблице 1. Таблица 1 - Группы сравнения по опытным дозировкам среды культивирования 100 Количество вещества, ммоль/л 25 6,25 1,56 0,39 FeSO4-7Н2О 1.1 1.2 1.3 1.4 1.5 Fe - nano 2.1 2.2 2.3 2.4 2.5 Fe - micro 3.1 3.2 3.3 3.4 3.5 CuSO4·5Н2О 4.1 4.2 4.3 4.4 4.5 Cu - nano 5.1 5.2 5.3 5.4 5.5 Cu - micro 6.1 6.2 6.3 6.4 6.5 Вещество Условия тестирования. Биотестирование проводили в лабораторных условиях в соответствии с ГОСТ 15150. Температура окружающего воздуха в лаборатории от + 18 до 25°С. Относительная влажность воздуха 80 ± 5%. Атмосферное давление 84-106 кПа (630-800 мм рт.ст.). Температура для 43 биотестирования (20+2)ºС. Освещение помещения естественное или искусственное, не ограничивается особыми требованиями. Термостаты перед проведением исследований промывали горячей водой с моющими средствами и дезинфицировали 1%-ным раствором марганцевокислого калия. На песок, в каждый контейнер, помещали по 30 семян пшеницы, которые заглубляли на глубину 5 мм. Сначала проводили посев контрольных семян, затем опытных с минимальными интервалами по времени. Контейнеры нумеровали. Подготовленные таким образом контейнеры помещали в термостат на проращивание. Условия проращивания: - температура постоянная + 20±2°С; - отсутствие освещенности; - влажность воздуха постоянная 80 ± 5%.; - экспозиция 14 дней. В случае необходимости проводили дополнительное орошение кварцевого песка дистиллированной водой при помощи дозатора (3-5 мл) на 5 и 10 сутки. По истечении времени экспозиции контейнеры извлекали из термостата и проводили необходимые наблюдения, исследования и замеры с определением: - показателей всхожести; - морфологических параметров корней и проростков; - аномалий развития. Биометрические исследования осуществляли общепринятыми методами, выполняя прямые измерения, а также фотографирование через лупу «Микромед» МС-1 (Россия-Китай) с последующим проведением измерений на полученных изображениях. После измерений образцы корневой зоны и зеленой части ростков пшеницы были герметично упакованы и временно заморожены для дальнейшего электронно-микроскопического исследования и определения микроэлементного состава. 44 Оценку эффектов наночастиц Cu и Fe на модели Allium сера проводили в двух вариантах. Первый (модифицированный) вариант Allium-теста в своей основе повторял описанную выше последовательность действий. В качестве субстрата использовали кварцевый песок (20 г), котрый помещали в преварительно стерилизованные в сушильном шкафу при температуре 130ºС в течение одного часа опытные контейнеры. Внесение исследуемого образца (FeSO4, CuSO4; наноматериалов Fe, Cu и микрочастиц Fe, Cu) в инертный субстрат осуществляли после диспергирования предварительно подготовленной навески в дистиллированной воде в течение 30 мин при 10 КГц. Максимальные дозировки (100 ммоль/л) подвергались многократному разведению. Подготовленными подобным образом растворами солей или лиозолей Cu и Fe песок увлажняли до полной влагоемкости, определенной по ГОСТ 12038-84. В контрольные контейнеры добавлялялись аналогичные объемы дистиллированной воды. На песок, в каждый контейнер, помещали по 2 луковицы массой 10-20 г и диаметром 1,5-2 см, которые заглубляли на 5 мм. Сначала проводили посев контрольных луковиц, затем опытных с минимальными интервалами по времени. Подготовленные, таким образом, контейнеры помещали в термостат на проращивание (температура постоянная +20±2°С; отсутствие освещенности; влажность воздуха постоянная 80±5%; экспозиция 7 дней). По истечении времени экспозиции контейнеры извлекали из термостата, после чего, оценивали морфологические признаки корневой системы. Концевые отрезки корней отделяли и использовали для оценки митотической активности. При оценке влияния наночастиц на целостность ДНК растительных клеток использовали классический вариант Allium-теста. В данной серии экспериментов проращивание корней Allium cepa первоначально проводили в дистиллированной воде, после чего в течение 3 часов контактировали с водными суспензиями наночастиц Cu в широком диапазоне использованных концентраций (как указано в таблице 1). 45 2.2.1.2 Методы микроскопического исследования растительных организмов Перед проведением электронно-микроскопических исследований концевые отрезки корней Triticum aestivum были отмыты в проточной воде и зафиксированы в 2,5 % глутаровом альдегиде, приготовленном на натрийфосфатном буфере (0,1М, pH 7,2), в течение 12 часов. Образцы были трекратно промыты тем же буфером с добавлением сахарозы (6,8%) с выдержкой по 10-15 мин. после чего пост-фиксированы 1 % OsO4 в течение 3-4 часов при комнатной температуре. Дегидратацию образцов проводили в следующих восходящих концентрациях этанола: В 30% - ополаскивание несколько раз; В 40% - выдерживали 2 раза по 10 мин; В 50% - 2 раза по 10 мин; В 60% - 2 раза по 10 мин; В 70% - 2 раза по 10 мин; В 80% - 2 раза по 10 мин; В 90% - 2 раза по 10 мин; В 96% - 2 раза по 10 мин; В ацетоне -3 раза по 20 мин. Далее образцы выдерживали в окиси пропилена 2 раза по 30 минут, после чего проводили пропитывание смолой и окисью пропилена (ОП) по 24 часа в следующих соотношениях: 2:1 (ОП-смола) - 4:2 мл 2 часа; 1:1 (ОП-смола) - 4:4мл ночь; 1:2 (ОП-смола) - 2:4мл день. Далее образцы помещали в капсулы Бима и ставили на полимеризацию: сутки при 37, сутки при 45, сутки при 57 ºС. Срезы толщиной 50-70 нм получали на ультромикротоме «Reichert-Jung» (Германия) срезы, контрастировали в течении 20 минут в 2% водном растворе 46 уранилацетата и дополнительно окрашивали цитратом свинца по Рейнольдсу при 20ºС в течение 6 мин. Срезы просматривали на электронном микроскопе JEM-100 (Япония) при ускоряющем напряжении 80 кВ и инструментальном увеличении ×15000. Для световой микроскопии концевые фрагменты корней Allium cepa длиной 1 см были помещены во флакончики и зафиксированы в растворе (1 мл) этилового спирта и уксусной кислоты (фиксатор Кларка) в течение суток. Затем материал несколько раз промывали 70% спиртом, до исчезновения запаха, и переносили во флакончики с 70% этиловым спиртом (по 1 мл) для долговременного хранения проб. Для приготовления давленных временных препаратов корневых меристем: - корешки отмывали от спирта в дистиллированнной воде; - переносили в небольшие фарфоровые тигли с держателем, которые на 2/3 предварительно заполняли красителем (2 % ацетокармином); - тигель с материалом, доводили до кипения (с помощью спиртовки), после чего его оставляли на 5-10 минут для прокрашивания хромосом; - от окрашенного корешка лезвием отрезали кончик меристемы длинной 2-3 мм, помещали на предметное стекло в каплю 45% уксусной кислоты, накрывали покровным стеклом и аккуратно раздавливали до получения клеточного монослоя. Временные давленные препараты просматривали и фотографировали при увеличении ×400 на тринокулярном микроскопе «Альтами 136Т» (Россия) с цифровой окулярной камерой 8 МП, после чего на основании учета не менее 1000 клеток осуществляли расчет митотического индекса, а также оценивали морфологию клеточных ядер. 2.2.1.3 Метод оценки накопления и распределения меди и железа в тканях растительных организмов Количественный анализ поступления и распределения меди и железа в проростках Triticum aestivum был выполнен с использованием метода методом 47 атомно-абсорбционной спектрометрии на базе Центра коллективного ползования «Институт микро- и нанотехнологий Оренбургского государственного университета». На предварительном этапе навески корней и побегов высушивали до постоянной массы, после чего проводили пробоподготовку с использованием концентрированной азотной, серной и соляной кислот. К каждой навеске добавляли по 1 мл концентрированной азотной кислоты, выпаривали на электрической плитке до влажных солей; осадок растворяли в 0,5 мл воды, добавляли 0,5 мл серной кислоты и 0,5 мл соляной кислоты, кипятили на электрической плитке по 20-30 мин; охлаждали, доводили до 1 мл раствором 3%ой азотной кислоты. Количественное определение содержания меди и железа проводили с использованием атомно-абсорбционного спектрометра с электротермической атомизацией «Люмекс» МГА-915 (Россия), настройка и градуировка которого проводилась в с использованием стандартов и в соответствии с руководством по эксплуатации. Определение содержания железа и меди в анализируемых пробах проводили при следующих условиях (таблица 2). Таблица 2 – Условия проведения анализа меди и железа методом атомноабсорбционной спектрометрии Элемент Длина волны, нм Сушка пробы, °С / время сушки, с Температура атомизации, °С Время атомизации, с Fe Cu 248,3 324,7 110 / 40 110 / 40 2400 2200 1,7 1,5 Результаты измерения (Х) были представлены в виде: Х ± Δ (мкг), где Δ показатель точности методики и в дальнейшем использованы для расчета показателей накопления в пересчете на сухую биомассу образца (мкг/г). 48 2.2.1.4 Метод анализа ДНК растительных организмов Выделение ДНК из концевых отрезков корней Allium cepa проводили с использованием набора «ДНК-экстран-4» («Синтол», Россия). Необходимое оборудование и материалы: - центрифуга для пробирок 1,5 мл (10000g); - термостат для 1,5 мл пробирок; - дозаторы (от 1 до 1000 мкл); - морозильная камера (-20° C). Навески 20-30 мг исследуемого материала (концевых отрезков корешков лука) измельчали, помещали в 1,5 мл пробирку, после чего добавляли 500 мкл лизис-буфера и 10 мкл раствора РНКазы, инкубировали при +37ºС. В пробу добавляли 150 мкл сорбента (перед использованием сорбент встряхивали, т.к. возможно его оседание при хранении) и интенсивно переворачивали пробирку 4-5 раз, чтобы перемешать лизат с сорбентом. Образец центрифугировали 2 мин. при 10000 – 13000 об./мин., супернатант переносили в новую пробирку (сорбент должен остаться на дне и выбрасывается вместе с пробиркой). К супернатанту добавляли 150 мкл солевого буфера, 2-3 раза переворачивали пробирку, и помещали в морозильную камеру (-20° C) на 5 мин. Образец центрифугировали 7 мин при 10000 – 13000 об./мин. для осаждения протеинов, после чего супернатант переносили в новую пробирку. К вновь полученному супернатанту добавляли 250 мкл осаждающего буфера, перемешивали, и центрифугировали 5 мин при 13000 об./мин; буфер сливали. К сформировавшемуся осадку добавляли 600 мкл 70% раствора этанола, центрифугировали 1 мин при 13000 об./мин; спирт сливали. Полученный осадок ДНК сушили (1 мин при 56° C и 1 мин на воздухе), растворяли в 50-100 мкл воды или ТЕ-буфере. Электрофоретическое разделение выделенных молекул проводили в 0.8% агарозном геле в присутствии 0, 5 мкг/мл этидиум бромида при силе тока 100 мА и напряженности электрического поля 5 В/см, используя в качестве контроля HindIII-рестрикты ДНК фага λ длиной от 250 до 10000 н.п. («СибЭнзим», Россия). 49 Результат миграции ДНК оценивали на УФ-трансиллюминаторе «Vilber Lourmat» (Франция) при 254 нм, а полученные цифровые изображения обрабатывали с использованием программы «ImageJ». 2.2.2 Методы исследования с использованием люминесцирующих бактерий 2.2.2.1 Метод оценки токсичности соединений меди и железа в тесте тушения бактериальной биолюминесценции При планировании и выполнении исследований были использованы методические указания: МУ 1.2.2634-10 «Микробиологическая и молекулярно генетическая оценка воздействия наноматериалов на представителей микробиоценоза». – М.: Федеральный центр гигиены и эпидемиологии Роспотребнадзора, 2010. В качестве инструмента исследования была использована лиофилизированная культура микроорганизмов «Эколюм» по ТУ 2639-23600209792-01 (НВО «Иммунотех», Россия), представляющая собой сенсорный штамм Escherichia coli K12 TG1, конститутивно экспрессирующий luxCDABEгены морской бактерии Photobacterium leiognathi 54D10. Оборудование, материалы, реактивы: - измерительный блок люминометра LM-01Т («Immunotech», Чехия); - весы электронные РВ 214 по ГОСТ 24104-2001; - рН-метр 150 МИ по ГОСТ 50.2.036-2004; - термостат суховоздушный ТС-1/80 по ТУ 9452-002-00141798-97; - термометр лабораторный 0-550С, цена деления шкалы – 0,50С по ГОСТ 215-73; - сушильный электрический шкаф по ГОСТ 13474-79; - холодильник бытовой, обеспечивающий замораживание (-18 ± 1°С) и хранение проб (+2 -+4°С) по ГОСТ 26678-85; - часы сигнальные по ТУ 25-07-57; 50 - дозаторы с переменным объёмом дозирования 0,02-0,5 см3 ± 1,0% по ТУ 9452-002-33189998-2002; - цилиндры вместимостью 25, 50 см3 второго класса точности по ГОСТ 1770-74; - пипетки объемом 0,5 и 1,0 см3 по ГОСТ 29227-91; - стаканы стеклянные лабораторные вместимостью 10 и 50 см3 по ГОСТ 25336-82; - воронки лабораторные по ГОСТ 25336-82; - стаканчики для взвешивания (бюксы) диаметром 30 и 40 мм по ГОСТ 25336-82; - бумажные фильтры обеззоленные типа ФОБ (красная, белая ленты) по ТУ 6-09-1678-95; - флаконы и банки стеклянные с навинчивающейся крышкой или с притертой пробкой для хранения проб и реактивов вместимостью 10, 50, 100 см3; - подставка (из пластика, дерева) с углублением для пенициллиновых пузырьков или измерительных кювет, на которой можно разместить не менее 12 кювет; - вода дистиллированная по ГОСТ 6709-72. Согласно инструкции, биосенсор восстанавливали из лиофилизированного состояния охлажденной дистиллированной водой для теста на острую токсичность. В отдельной серии экспериментов для теста на хроническую токсичность дистиллированную воду заменяли равным объемом LB-бульона. Сравнительное изучение токсичности ионов, микро- и наночастиц меди и железа проводилось в тесте ингибирования бактериальной биолюминесценции, отражающем изменение энергетики бактериальной клетки при подобном воздействии. Для этого эквимолярные количества предварительно подготовленных растворов и суспензий в объеме 100 мкл вносили в ячейки 96луночного планшета из непрозрачного пластика, где готовили их последовательные разведения от 1:2 до 1:1024. После этого в каждую ячейку вносили по 100 мкл биосенсора, после чего планшет помещали в измерительный 51 блок люминометра LM-01Т («Immunotech», Чехия), динамически регистрируя интенсивность свечения через 1, 3, 6 и 16 ч. Выраженность ингибирования бактериальной биолюминесценции (Iинг) оценивали по формуле , где Ik и Io - интенсивность свечения контрольных и опытных проб при фиксированном времени экспозиции исследуемого раствора или суспензии металла с тест-объектом. Полученные результаты использовали для расчета величин токсикологического параметра ЕС50, соответствующих молярным концентрациям исследуемых соединений железа и меди, вызывающих подавление бактериального свечения на 50 %, по сравнению с контролем. 2.2.2.2 Метод оценки механизмов биоактивности соединений меди и железа в тесте индукции бактериальной биолюминесценции Исследование способности соединений меди к индукции окислительного стресса и связанного с этим окислительного повреждения ДНК бактериальных клеток-мишеней было проведено на основе определения индукции свечения репортерных штаммов на основе E.coli K12 MG1655, несущих плазмиды с генными слияниями soxS::lux, katG::lux, recA::lux Перед проведением исследований микроорганизмы выращивали в LBбульоне до достижения ранней фазы логарифмического роста, после чего биомассу отмывали центрифугированием и ресуспендировали в растворе 0.85 % NaCl. В ячейках 96-луночного планшета из непрозрачного пластика готовили последовательные разведения растворов или суспензий анализируемых соединений меди от 1:2 до 1:1024 как описано в разделе 2.2.2.1, после чего в каждую ячейку вносили равные объемы бактериальной культуры. Полученную смесь инкубировали в течение 15 минут для проявления биоактивности соединений меди, после чего в ячейки дополнительно вносили по 52 100 мкл LB-бульона в качестве условия для проявления индукции биолюминесценции. Планшет помещали в измерительный блок люминометра, где в течение 120 мин динамически регистрировали развитие бактериального свечения. Уровень экспрессии генов-репортеров, отражающий индукцию соответствующих промоторов (Iинд), оценивали по приведенной выше формуле. 2.2.3 Статистическая обработка результатов Эксперименты выполнены не менее чем в трех повторностях. Обработку полученных данных осуществляли с помощью компьютерной программы «Еxcel 2010» (Microsoft Inc., 2003), рассчитывая среднее арифметическое и стандартную ошибку. Достоверность различий оценивалась с использованием критерия Стьюдента. 53 ГЛАВА 3. СРАВНИТЕЛЬНАЯ ОЦЕНКА ТОЛЕРАНТНОСТИ РАСТЕНИЙ (В ТЕСТЕ ПРОРАСТАНИЯ СЕМЯН TRITICUM AESTIVUM) И МИКРООРГАНИЗМОВ (В ТЕСТЕ ИНГИБИРОВАНИЯ БИОЛЮМИНЕСЦЕНЦИИ ESHERICHIA COLI) К ВОЗДЕЙСТВИЮ НАНОЧАСТИЦ, МИКРОЧАСТИЦ И СОЛЕЙ ЖЕЛЕЗА И МЕДИ 3.1. Оценка безопасности ионов, нано- и микрочастиц Fe и Cu в тесте прорастания семян Triticum aestivum Результаты сравнительного изучения биологической активности наночастиц Fe и Cu с ионами и микрочастицами данных металлов в тесте прорастания семян пшеницы Triticum aestivum, рекомендуемом для заявленных целей действующим национальным нормативом (Методические указания 1.2.2635-10 Медико – биологическая оценка безопасности наноматериалов. М.: Федеральный центр гигиены и эпидемиологии Роспотребнадзора. 2010. 123 с.), позволили продемонстрировать выраженные зависимости регистрируемых проявлений фитотоксичности от природы исследуемых металлов, их действующих форм и концентраций. Так инкубация семян Triticum aestivum в контакте с ионами железа в концентрации 100 ммоль/л полностью подавляла их пророст, в концентрации 25 ммоль/л снижала частоту прорастания до 13,3±6,8% против 98,3±2,6% в контроле (P<0,001), в то время как дальнейшее снижение присутствия данного фактора полностью снимало указанный эффект. Более выраженные изменения были констатированы при анализе размерных характеристик побегов и корней проростков. Так концентрация ионов железа 25 ммоль/л полностью блокировала развитие корневой системы, одновременно почти десятикратно сокращая длину побега: до 7,0±1,1 мм против 66,3±4,6 мм в контроле (P<0,05). В свою очередь концентрация ионов железа 6,25ммоль/л снижала длину корней до 16,0±0,1 мм против 221,4±21,3 мм, а длину побега в два раза по сравнению с соответствующим контролем: до 38,6±2,5 мм (P<0,05). Рассчитанные на данной 54 основе значения EC50 (таблица 3) соответствующие подавлению результативного параметра на 50% от контрольных значений и тем самым характеризующие верхний предел толерантности к ионам железа, свидетельствовали об увеличении выраженности токсического эффекта в ряду «всхожесть семян (17 ммоль/л) → длина побега (6,3 ммоль/л) → длина корня (3,3 ммоль/л)». Таблица 3 - Значения верхнего предела толерантности (параметр ЕС50), определенные в тесте прорастания семян Triticum aestivum в среде, содержащей различные формы Fe и Cu (ммоль/л). Параметры эффекта подавление уменьшение уменьшение всхожести семян длины побега длины корня 17,0 6,3 3,3 Металлы и их исследуемые формы ионы Fe наночастицы >100 >100 >100 микрочастицы >100 >100 >100 4,0 3,5 0,2 наночастицы >100 >100 1,7 микрочастицы >100 >100 6,7 ионы Cu На этом фоне использование идентичных концентраций нано- и микрочастиц Fe достоверно не сказывалось на частоте прорастания семян, в подобных случаях составившей 90,0-93,3% и 96,7-100,0% соответственно. Сходный результат получен и при исследовании размерных характеристик побегов и корней, в используемом диапазоне концентраций даже имеющих некоторую тенденцию к увеличению. Сказанное позволило констатировать высокий уровень толерантности Triticum aestivum к железу в нано- и микроструктурированной формах, что нашло свое отражение при расчете соответствующих величин EC50 (таблица 3), а также хорошо согласовывалось с известными данными по этому вопросу. В частности, выявленная при обработке нано- и микрочастицами Fe умеренная стимуляция параметров прорастания Triticum aestivum подтверждает целесообразность проведения предпосевной 55 обработки семян данными соединениями, в оптимальных дозах дающей от 5 до 30% повышения урожайности и улучшения товарного вида растительной продукции (Коваленко Л.В., Фолманис Г.Э. Биологически активные нанопорошки железа. М.: Наука, 2006. 124с.). Проведение аналогичных исследований с использованием нано-, микрочастиц и ионов меди свидетельствовало о более низкой толерантности Triticum aestivum к воздействию этого металла (рисунок 2). Рисунок 2 - Эффекты ионов, нано- и микрочастиц меди в концентрации 100 ммоль/л на модели проростков Triticum aestivum 56 В частности, анализ морфологии проростков Triticum aestivum, инкубированных в присутствии наночастиц Cu, позволил констатировать статистически значимое изменение ряда биометрических параметров, преимущественно относящихся к корневой системе данного растения. Так наночастицы меди хотя достоверно и не изменяли частоту прорастания семян, но в максимальной использованной концентрации 100 ммоль/л негативно влияли на длину побегов, а наиболее выражено подавляли формирование корневой системы (EC50=1,7 ммоль/л). На этом фоне еще меньшая толерантность была зарегистрирована к ионной форме меди, выражено подавляющей частоту всхожести семян Triticum aestivum (EC50=4 ммоль/л), а также достоверно уменьшающей длину формирующихся побегов (EC50=3,5 ммоль/л) и корней (EC50=0,2 ммоль/л), что в 2-4-16 раз превышало аналогичные значения для ионов железа. В свою очередь микрочастицы Cu, не оказывающие достоверного угнетающего влияния на частоту прорастания семян и длину побегов, все-таки подавляли формирование корневой системы, что характеризовалось значением EC50=6,7 ммоль/л (таблица 3). Таким образом, совокупность полученных результатов, с одной стороны, позволили четко продемонстрировать подтвердить закономерность увеличения фитотоксического эффекта металлов в ряду «всхожесть семян → длина побега → длина корня)», а с другой – при анализе последнего параметра констатировать рост подобной активности соединений меди в ряду «микрочастицы → наночастицы → ионы». Сопоставление полученных результатов с известными данными по этому вопросу хорошо корреспондируются с недавними наблюдениями группы исследователей из университета Нью-Хейвена (Коннектикут, США), также указывающими на выраженную фитотоксичность наночастиц меди, сопоставимую с таковой у соединений серебра (Stampoulis D., Sinha S. K., White J.C. Assay-dependent phytotoxicity of nanoparticles to plants // Environ. Sci. Technol. 57 2009, Vol. 43, P. 9473−9479, Musante C., White J.C. Toxicity of silver and copper to Cucurbita pepo: differential effects of nano and bulk-size particles // Environ. Toxicol. 2012, Vol. 27. P. 510-517). Одновременно накапливается все больше экспериментальных данных о том, что основной «мишенью» для наночастиц металлов является именно корневая система растений, что диктует интерес к выявлению механизмов подобной фитотоксичности, в том числе с акцентом на исследование митотических процессов в меристеме, определяющих рост корня в длину, что определило задачу продолжения поиска в заданном направлении. 3.2. Биологическая активность ионов, нано- и микрочастиц Cu и Fe в тесте ингибирования бактериальной биолюминесценции Исследование пределов толерантности живых объектов к воздействию соединений железа бактериальной и меди было биолюминесценции продолжено в рекомбинантного тесте ингибирования люминесцирующего штамма Escherichia coli, также рекомендуемого для медико-биологической оценки наноматериалов действующим национальным нормативом (МУК 1.2. 2634-10). При этом совокупность полученных данных позволила более полно охарактеризовать динамику развивающегося эффекта, а также подтвердить его зависимость от природы исследуемых металлов, их действующих форм и концентраций. Так контакт E. coli с возрастающими концентрациями ионов меди в растворе соли в диапазоне от 100 до 0,78 ммоль/л вел к развивающемуся в первые 15-60 минут выраженному токсическому эффекту - полному подавлению бактериального свечения (рисунок 3), что могло интерпретироваться как проявление выраженной острой токсичности данного вещества. В свою очередь для формирования аналогичного эффекта ионов меди в концентрации 0,39 ммоль/л требовалось уже 170 мин, а еще более низкие дозировки обуславливали лишь относительную ингибицию бактериальной биолюминесценции до 10-70 % от контрольных значений. Аналогичный результат зафиксирован и при тестировании возрастающих концентраций ионов Fe2+, по своим динамическим и количественным 58 характеристикам достоверно не отличающийся от такового у ионов меди. Выраженная активность ионов железа в использованной биоиндикационной системе могла объясняться ранее показанной пероксидацией мембранных липидов E. coli в присутствии металлов переменной валентности, продукты которой способны выступать в качестве ингибиторов ферментной системы генерации свечения. Lg (I) 10000 Lg (I) 10000 А Lg (I) 10000 Б В 1000 1000 1000 100 100 100 10 10 10 1 1 0 20 40 60 80 100 120 140 160 180 Время, мин. 1 0 20 40 60 80 100 120 140 160 180 Время, мин. 0 20 40 60 80 100 120 140 160 180 Время, мин. Рисунок 3 - Динамика свечения E. coli K12 TG1 с клонированными luxCDABE-генами P. leiognathi при контакте с ионами (А), нано- (Б) и микрочастицами (В) меди, использованными в концентрациях 100 (1); 50 (2); 25 (3); 12,5 (4); 6,25 (5); 3,12 (6); 1,56 (7); 0,78 (8); 0,39 (9); 0,19 (10) и 0,09 (11) ммоль/л; контроль (к). По оси абсцисс – время контакта, мин; по оси ординат – регистрируемые значения интенсивности свечения. На этом фоне наночастицы меди также обуславливали ингибирование бактериальной биолюминесценции, количественные характеристики которого, однако, имели ряд существенных отличий. Первым из них было замедленное формирование токсического эффекта, развивающегося только со 120-й минуты контакта. Второй особенностью стало то, что для достижения подобного эффекта 59 требовались значительно более высокие концентрации наночастиц меди - от 12,5 до 100 ммоль/л (рисунок 3б) по сравнению с ионной формой данного металла. Еще в большей степени толерантность бактерий была выражена в отношении наночастиц железа, в стандартных условиях не проявляющих токсического эффекта, а в отдельной серии экспериментов, предусматривающей выращивание E. coli в среде с тестируемыми соединениями, все-таки вызывающих подавление бактериального свечения, но только в максимальной использованной концентрации и только при высокой продолжительности воздействия. На этом фоне микрочастицы Cu и Fe проявляли себя как наиболее биологически инертные образцы, среди которых способность к ингибированию бактериальной биолюминесценции проявляли только микрочастицы меди в максимальной использованной концентрации 100 ммоль/л (рисунок 3в). Приведенные данные явились основой для построения зависимостей «дозаэффект» для каждого из исследуемых соединений (рисунок 4). 1,8 I А 1,8 1,6 1,6 1,4 1,4 1,2 1,2 1 1 0,8 0,8 0,6 0,6 0,4 0,4 0,2 0,2 0 0 0 Рисунок 20 40 4 - 60 80 100 Концентрация, мМ I Б 0 Относительные 20 40 величины 60 80 100 Концентрация, мМ интенсивности свечения люминесцирующего штамма E. coli при контакте с ионами (1), нано- (2) и микрочастицами (3) меди (А) и железа (Б). 60 Расчет величин ЕС50, отражающих верхний предел толерантности бактерий и соответствующих молярным концентрациям Cu и Fe, вызывающим 50% ингибирование бактериальной биолюминесценции по сравнению с контролем при различной продолжительности воздействия представлен в таблице 4. Таблица 4 - Значения верхнего предела толерантности (параметр ЕС50) рекомбинантного штамма E. coli K12 TG1 с клонированными luxCDABE-генами P.leiognathi к воздействию различных форм Fe и Cu (ммоль/л). Металлы и их исследуемые формы Fe Cu Продолжительность воздействия, мин ионы 60 0,070 120 0,065 180 0,060 наночастицы >100 >100 >100 микрочастицы >100 >100 >100 ионы 0,083 0,068 0,058 наночастицы >100 58,0 1,5 микрочастицы >100 96,0 75,0 Проведение подобных вычислений позволило констатировать, что характеризуемый величинами ЕС50 уровень токсичности исследуемых соединений металлов в отношении рекомбинантного люминесцирующего штамма E. coli прогрессивно убывал в ряду «ионы → наночастицы → микрочастицы», а сам подобный эффект в наибольшей степени оказывался выраженным у соединений меди по сравнению с соединениями железа. Так особенностью биологической активности ионов Cu2+ являлось формирование значимых величин ЕС50 уже к 60-й минуте контакта, незначительно изменяющихся до продолжительности 0,058±0,0003 воздействия. ммоль/л На при этом фоне дальнейшем увеличении использование нано- и микрочастиц меди существенно снижало скорость и выраженность токсических эффектов, становящихся значимыми только со 120-й минуты контакта, а к 180-й 61 минуте более чем в 30 раз (ЕС50 = 1,5±0,0075 ммоль/л) и 1500 раз (ЕС50 = 75±3,5 ммоль/л) уступающих аналогичной активностью данного металла в ионной форме. Одновременно полученные данные хорошо согласовывались с приведенными выше результатами тестирования данного металла на Triticum aestivum, а также данными Z. Chen с соавт., зафиксировавшими выраженное снижение токсичности нано- и микрочастиц меди, характеризуемой значениями LD50 для мышей 413 и >5000 мг/кг, против LD50 = 110 мг/кг для CuCl2. С другой стороны, при общем совпадении обозначенных тенденций абсолютные значения токсичности одних и тех же соединений меди для растений оказались выше, чем для микроорганизмов, что подтверждает увеличение толерантности живых систем с ростом уровня сложности их организации. На этом фоне особенностью исследованных соединений железа явилась способность к ингибиции бактериальной биолюминесценции только у его ионной формы, вновь развивающейся уже на ранних сроках воздействия и в дальнейшем изменяющейся до 0,06±0,0003 ммоль/л. В свою очередь использование технологии биолюминесцентного биотестирования не позволило зафиксировать токсических свойств как у нано-, так и у микрочастиц Fe, что подтверждает представления о высоком уровне биосовместимости данных соединений. Те же данные позволяют по-разному оценивать последствия поступления наночастиц Cu и Fe в абиотические среды и компоненты биоты, характеризуя первые из них как экологически безопасные соединения, а для других требуя обоснования и утверждения соответствующих санитарно-гигиенических нормативов и предельно-допустимых концентраций. Таким образом, результаты первого этапа исследований позволили получить развернутые представления о значениях верхнего предела толерантности модельных растений и бактерий к препаратам железа и меди, закономерно снижающейся в ряду «микрочастицы → наночастицы → ионы» и нарастающей от меди к железу. При этом детали подобного эффекта заключались в определении корневой системы растений как основной «мишени» для воздействия металлических наночастиц, формирующем основу для продолжения 62 исследований в заданном направлении для выявления механизмов регистрируемого фитотоксического эффекта. Кроме того, микроорганизмы оказывались более чувствительными к действию исследуемых соединений металлов чем растения, однако, существенный параллелелизм результатов исследования биологической активности ионов, нано- и микрочастиц в растительном биотесте и тесте ингибирования бактериальной биолюминесценции определил возможность последующего использования данных о тонких механизмах токсичности названных соединений, получаемых с использованием бактериальных моделей, и для объяснения характера их действия в отношении высших растений. 63 ГЛАВА 4. АДАПТИВНЫЕ РЕАКЦИИ РАСТЕНИЙ К ПРИСУТСТВИЮ В СРЕДЕ КУЛЬТИВИРОВАНИЯ НАНОЧАСТИЦ МЕДИ 4.1. Анализ механизмов биологической активности наночастиц меди в тестах на Triticum aestivum и Allium cepa Полученные результаты, свидетельствующие о низкой толерантности растений к воздействию наночастиц меди, определили перспективу продолжения исследований в направлении идентификации тонких механизмов их биологической активности, в том числе закономерностей поступления, распределения и накопления в растительных тканях, а также природы развивающегося при этом токсического эффекта. При этом для решения этой задачи был привлечен комплекс методов биометрического, микроскопического, химического и генетического анализа, реализованных при оценке эффектов наночастиц меди в отношении Triticum aestivum и Allium cepa. Развернутый анализ морфологии проростков Triticum aestivum, инкубированных в присутствии различных концентраций наночастиц Cu, позволил детализировать представления о статистически значимом изменении ряда биометрических параметров данного модельного растения, преимущественно относящихся к его корневой системе. Так наночастицы меди достоверно не изменяли частоту прорастания семян, негативно влияли на массу и длину побегов только в максимальной использованной концентрации 0,1 М, но выражено подавляли формирование корневой системы, что проявлялось в уменьшении средней длины и массы образующихся корней в широком диапазоне воздействующих концентраций наночастиц Cu (таблица 5). В частности, присутствие в водной фазе среды культивирования наночастиц меди в концентрациях 0,025 - 0,1 М полностью блокировало развитие корневой системы, а в концентрации 6,25 мМ уменьшало длину корней до 38,3±4,1 мм (P<0,001) и их массу до 18,7±2,5 мг (P<0,001) против аналогичных значений 221,4±21,3 мм и 58,5±3,1 мг в контроле, что сохранялось на статистически значимом уровне вплоть до концентрации 1,56 мМ. 64 Таблица 5 - Характеристика корневой системы Triticum aestivum при культивировании в среде, содержащей различные концентрации наночастиц меди Концентрация наночастиц, ммоль/л Длина первичного корня, мм Количество Длина придаточных придаточных корней, шт. корней, мм Контроль 98,9±5,54 2,13±0,06 76,5±4,12 100 2,46±0,61*** 3,7±0,34* 2,25±0,18*** 25 3,6±1,6*** 4,2±0,2* 2,41±0,22*** 6,25 15,1±2,34** 4,1±0,27* 5,69±0,66*** 1,56 45,9±4,54* 2,79±0,19 25,71±1,77* 0,39 95,9±9,08 2,26±0,16 71,77±4,82 Обозначения: * - P<0,05; ** - P<0,01; *** - P<0,001 Формализованным выражением подобного фитотоксического эффекта стали значения ЕС50 = 1,7 мМ (для длины корня) и ЕС50 = 2,8 мМ (для массы корня), соответствующие концентрациям наночастиц Cu, подавляющим указанные биометрические величины на 50% от контрольных значений. Помимо этого результатом воздействия наночастиц меди на Triticum aestivum являлось увеличение диаметра образующихся корней, а также их не регистрируемое в контрольных образцах ветвление по типу «куриной лапки» (рисунок 5). Тем самым наблюдениями полученные группы результаты исследователей хорошо из корреспондируются университета с Нью-Хейвена (Коннектикут, США), в своих работах (Musante C., White J.C. Toxicity of silver and copper to Cucurbita pepo: differential effects of nano and bulk-size particles // Environ. Toxicol. 2012. Vol. 27. P. 510-517) также указывающих на выраженную фитотоксичность наночастиц Cu. При этом накапливающиеся данные о биологической активности наночастиц металлов в растительных биотестах (Lee W. M., An Y. J., Yoon H., Kweon H.S. Toxicity and bioavailability of copper 65 Рисунок 5 - Морфология корневой системы Triticum aestivum в контроле (слева) и при проращивании в среде с содержанием 6,25 мМ наночастиц Cu (справа); нижний ряд - увеличение × 8. nanoparticles to the terrestrial plants mung bean (Phaseolus radiatus) and wheat (Triticum aestivum): plant agar test for water-insoluble nanoparticles // Environ. Toxicol. Chem. 2008. Vol. 27. P. 1915−1921; Lee C.W., Mahendra S., Zodrow K. Developmental phytotoxicity of metal oxide nanoparticles to Arabidopsis thaliana // Environ. Toxicol. Chem. 2010. Vol. 29. P. 669-675) в качестве основной «мишени» их действия уверенно позволяют назвать именно корневую систему модельных растений. 66 На следующем этапе исследований отрезки контрольных и опытных образцов корней пшеницы, включающие апикальную часть (зону деления) вместе с корневым чехликом и первичной меристемой конуса нарастания (0-2 мм от кончика корня), расположенную непосредственно ниже зоны растяжения, были представлены для электронно-микроскопического исследования Цитологическая характеристика визуализированных в контроле клеток позволила констатировать у них сходные и характерные для меристемы черты ультраструктуры (рисунок 6 а, б). При этом исследованная зона корня состояла из плотно расположенных изодиаметрических клеток размером от 6×14 до 10×25 мкм с плотной цитоплазмой и относительно крупными ядрами диаметром от 1,5 до 2 мкм. В их цитоплазме обнаруживались свободные рибосомы и короткие профили эндоплазматического ретикулума, единичные пластиды, а также более многочисленные митохондрии длиной до 1,7 мкм и шириной около 0,25 мкм с характерной морфологией. Кроме того, в клетках присутствовал в различной степени развитый вакуолярный аппарат, представленный округлыми образованиями диаметром 100-200 нм с электронно-прозрачным содержимым. Клетки контрольных образцов корня были окружены плотно прилегающими клеточными стенками толщиной 246±45 нм, при этом использованный метод контрастирования не визуализировал в них заметного количества плазмодесм. В свою пшеницы, очередь, выращенной электронно-микроскопическое в присутствии исследование наночастиц меди, корней позволило констатировать ряд ультраструктурных изменений, в частности, затрагивающих характер организации клеточных стенок. При этом в них обнаруживались многочисленные плазмодесмы, в отсутствие дополнительного контрастирования визуализируемые как электронно-плотные полосы, пересекающих клеточные стенки на расстоянии 0,5-2 мкм друг от друга (рисунок 6 в, г). В использованном экспериментальном контексте формирование дополнительной электронной плотности на участках плазмодесм может быть следствием прохождения через них наночастиц, оставляющих за собой «следы» из диссоциировавших с поверхности кластеров меди. 67 Рисунок 6 - Фрагменты ультраструктуры клеток Triticum aestivum в контроле (а, б) и при проращивании в среде с содержанием 6,25 мМ наночастиц Cu (в, г). Обозначения: кс – клеточная стенка; я – ядро; мх – митохондрия; в – вакуоли; нч – наночастицы. Стрелкой указано место предполагаемого прохождения нанчастиц меди через плазмодесму. 68 Дополнительными аргументами в пользу данного предположения являются сопоставимость диаметра плазмодесм (до 70 нм) и использованных наночастиц (84±5 нм), а также регистируемая на некоторых электроннограммах ассоциированность последних с местами открытия плазмодесм в цитозоль растительных клеток (рисунок 6г). Кроме того, важной особенностью оказывалось более чем двукратное (684±120 нм, P<0,01) утолщение клеточных стенок в корнях, выросших в присутствии наночастиц меди. Данный факт, с одной стороны, может рассматриваться как одно из проявлений реакции тканей растения на проникновение и распространение наночастиц, а с другой – объяснять природу утолщения корней пшеницы при воздействии наночастиц Cu по сравнению с соответствующим контролем. Другим важным результатом электронно-микроскопического исследования являлась визуализация в цитоплазме клеток опытных образцов корня Triticum aestivum многочисленных электронно-плотных образований округлой формы, по своим характеристикам полностью соответствующих заявленным параметрам использованных наночастиц меди. Сказанное свидетельствует в пользу возможности поступления наночастиц Cu в ткани растений в их исходной наноструктурированной форме, где предполагаемой движущей силой данного процесса является градиент концентраций наночастиц в системе «среда культивирования – ткань корня», а механизмом распространения – межклеточный перенос вглубь корня через систему плазмодесм. Тем самым полученный результат хорошо согласуется с данными (Faisal M., Saquib Q., Alatar A.A., AlKhedhairy A.A., Hegazy A.K., Musarrat J. Phytotoxic hazards of NiO-nanoparticles in tomato: A study on mechanism of cell death // J. Hazard. Mater. 2013. Vol. 250-251. P. 318-332), на примере наночастиц NiO также демонстрирующими возможность распространения наночастиц вглубь корней томатов через систему межклеточных контактов и далее – по сосудистой системе растения. В продолжение анализа распределения наночастиц меди в тканях модельных растений количественное содержание этого элемента с пересчетом на сухую биомассу Triticum aestivum было исследовано методом атомно-адсорбционной 69 спектроскопии. При этом если присутствие меди в контрольных образцах описывалось величинами 4,78±1,57 мкг/г в корневой системе проростка и 5,19±1,71 мкг/г в его зеленом побеге, то инкубация в контакте с наночастицами дозозависимо статистически значимо присутствие этого элемента (таблица 6). Таблица 6 - Количественное содержание меди в проростках Triticum aestivum (в пересчете на сухую биомассу, мкг/г), инкубированных в присутствии различных концентраций наночастиц Cu Исследуемые Концентрация наночастиц меди части в среде культивирования, мкг/мл Triticum aestivum Побеги Корни 6350 1587,5 396,9 99,2 291,32 150,99 36,05 18,18 ±64,09 ±33,22 ±7,93 ±4,00 *** *** ** * 287,91 46,34 ±63,34 ±10,19 *** ** н/о н/о Контроль 24,8 7,86 5,19 ±2,59 ±1,71 6,91 4,78 ±2,28 ±1,57 Обозначения: * - P<0,05; ** - P<0,01; *** - P<0,001; н/о – не определяли Так проращивание семян пшеницы в среде с содержанием наночастиц Cu от 0,78 мМ (24,8 мкг/мл) до 6,25 мМ (396,9 мкг/мл) вело к выраженному накоплению меди в формирующейся корневой системе: от 6,91±2,28 до 287,91±63,34 мкг/г, что соответствовало 27,8-72,5 % эффективности извлечения данного металла из среды культивирования. В свою очередь присутствие меди в зеленых частях проростков Triticum aestivum во всем используемом диапазоне воздействующих концентраций возрастало от 7,86±2,59 до 291,32±64,09 мкг/г, что в сопоставимых пробах в 2,57,9 раза уступало ее содержанию в тканях корня и соответствовало 4,6-18,3 % эффективности извлечения из среды культивирования. Тем самым полученные результаты демонстрируют возможность пропорционального воздействующей концентрации наночастиц Cu поступления и распространения меди в тканях 70 проростков пшеницы с преимущественным накоплением в корневой системе, что развивает положение о последней как основной «мишени» для реализации биологической активности наночастиц. Исследование непосредственных механизмов фитотоксичности наночастиц Cu было проведено в модифицированном варианте Allium-теста, ранее с успехом использованного для анализа биологической активности наночастиц серебра (Babu K., Deepa M., Shankar S., Rai S. Effect of nano-silver on cell division and mitotic chromosomes: A prefatory siren // The Internet Journal of Nanotechnology. 2008. Vol. 2(2). P. 1−14) и диоксида титана (Klancnik K., Drobne D., Valant J., Dolenc K.J. Use of a modified Allium test with nanoTiO2 // Ecotoxicol. Environ. Saf. 2011. Vol. 74. P. 85-92). Проведенный анализ контрольных образцов позволил выявить типичную для меристематических тканей корня лука картину, характеризующуюся присутствием 7,48±1,10 % клеток, находящихся на различных стадиях митоза (рисунок 7а). В свою очередь проращивание луковиц Allium cepa в присутствии наночастиц меди позволило констатировать уменьшение средней длины образующихся корней (ЕС50 = 10,5 мМ), что в сравнении с аналогичными результатами на Triticum aestivum свидетельствовало о несколько меньшей чувствительности данного модельного растения к оцениваемому воздействию, но по своей направленности и выраженности хорошо согласовывалось с аналогичными результатами на проростках пшеницы. Помимо сдерживания корневого прироста макроскопический анализ опытных образцов корней лука фиксировал изменение цвета (потемнение) их концевых участков, что могло свидетельствовать как об интенсивном проникновении наночастиц в меристему, так и о развивающемся здесь цитотоксическом эффекте. В свою очередь микроскопическое исследование корней, пророщенных в контакте с наночастиц Cu свидетельствовало о полном подавлении пролиферативной активности визуализированных клеток со стремящимися к 71 Рисунок 7 - Митотоксическое действие наночастиц меди в модифицированном варианте Allium-теста: а – контроль; б – опыт (проращивание в присутствии 6,25 мМ наночастиц Cu). Обозначения: м – клетки, находящиеся на разных стадиях митоза; кпя – кариопикнотические ядра. нулю значениями митотического индекса, сохраняющем клетки в интерфазном состоянии и делающем технически невозможным регистрацию митоз- модифицирующих или мутагенных эффектов. Дополнительным проявлением подобного митотоксического эффекта наночастиц меди являлись деструкция и конденсация хроматина (рисунок 7б), что соответствует процессу пикнотического разрушения ядра и по существующим представлениям является одним из признаков гибели растительных клеток. 72 Аналогичная, но более детальная картина морфологических изменений клеток Triticum aestivum, выращенных в присутствии наночастиц Cu, была зарегистрирована при проведении электронной микроскопии (рисунок 8). А Б В Рисунок 8 - Изменения ультраструктуры клеток корня пшеницы Triticum aestivum при воздействии наночастиц меди: а – образование цитоплазматических вакуолей; б – конденсация хроматина в ядре; в – полное разрушение клеток корня, содержащих электронно-плотные наночастицы. 73 При этом в препаратах корней пшеницы, пророщенной в контакте с наночастицами меди, также обнаруживался ряд ультраструктурных проявлений клеточной гибели. Сказанное заключалось в существенной вакуолизации цитоплазмы: данные структуры увеличивались в размерах до 500-800 нм, а в их составе визуализировались остатки отдельных органелл, в т.ч. митохондрий (рисунок 8а). Интерес также представляла выявляемая в некоторых клетках конденсация ядерного хроматина с сопутствующим расширением перинуклеарного пространства и формированием инвагинаций (рисунок 8б), что соответствует описанному выше процессу пикнотического разрушения ядра. Наконец крайним проявлением происходяших событий являлось полное разрушение клеток корня, остатки которых были представлены окруженным клеточной стенкой бесструктурным матриксом, включающим электронноплотные образования, по своей форме и размеру соответствующие использованным наночастицам Cu (рисунок 8в). С целью выяснения природы регистрируемых явлений в отдельной серии экспериментов действие наночастиц меди было оценено в классическом варианте Allium-теста, подразумевающем контакт их суспензии с предварительно пророщенной корневой системой лука. При этом электрофоретическое разделение ДНК, выделенной из концевых отрезков корней лука после 3-часовой инкубации в среде с наночастицами меди, позволило констатировать существенную и дозозависимую деградацию данного биополимера (рисунок 9). Так, если электрофоретическая подвижность выделенной их контрольных образцов ДНК характеризовала ее как достаточно протяженные полимерные молекулы, основная доля которых представлена участками > 10 тысяч н.п., то результатом воздействия наночастиц Cu оказывалось существенное укорочение изолированных фрагментов ДНК. В частности, результатом воздействия максимальной (0,1 М) концентрации наночастиц меди являлась экстремально выраженная деградация ДНК с превращением 80 % данного биополимера в короткоцепочечные фрагменты < 1000 н.п. 74 Рисунок 9 - Электрофореграмма ДНК, выделенной из концевых отрезков корней Allium cepa, инкубированных в контакте с наночастицами меди (слева) и основанный на этом расчет соотношения длинноцепочечных и короткоцепочечных фрагментов ДНК (справа). Обозначения: м – маркеры электрофоретической подвижности ДНК, стрелкой указано положение маркера 1000 н.п.; к – контроль; 1,2,3 – корни, инкубированные в контакте с наночастицами меди в концентрациях 0,1; 0,025 и 0,00625 М, соответственно; темная часть столбчатой диаграммы – доля фрагментов размером >1000 н.п., светлая часть - доля фрагментов размером < 1000 н.п. Снижение воздействующей концентрации наноCu до 0,025 М сохраняло до двух третей ДНК в длинноцепочечной форме, в то время как 35 % по-прежнему имели размер < 1000 н.п. Наконец понижение присутствия наночастиц меди до 0,00625 М переводило в короткоцепочечную форму не более 10 % изолированных фрагментов ДНК. Отдельным важным аспектом полученного результата является регистрация деградированной ДНК в виде единого трека, но не «лестницы» фрагментов (как следствия межнуклеосомальной ферментативной фрагментации данного биополимера), что является характерным для гибели растительных клеток по пути некроза, но не апоптоза (Ванюшин Б.Ф. Апоптоз у растений // Успехи биологической химии. 2001. Т. 41. С. 3-38). 75 Ранее индуцируемое наночастицами меди повреждение ДНК было показано на проростках редиса (Raphanus sativus) и райграса (Lolium perenne и Lolium rigidum), что сопровождалось накоплением в данном биополимере окисленных нуклеотидов: 7,8-дигидро-8-оксогуанина; 2,6-диаминор-4-гидрокси-5- формамидопиримидина и 4,6-диамино-5- формамидопиримидина (Atha D.H., Wang H., Petersen E.J., Cleveland D., Holbrook R.D., Jaruga P., Dizdaroglu M., Xing B., Nelson B.C. Copper oxide nanoparticle mediated DNA damage in terrestrial plant models // Environ. Sci. Technol. 2012. Vol. 46. P. 1819−1827) и также вело к существенному подавлению роста модельных растений. Конечным же следствием происходящих событий является остановка растительных клеток в одной из точек контроля клеточного цикла (наиболее вероятно G1-S), где обнаруживающий поврежденную ДНК механизм прекращает процесс деления. Таким образом, проведенное исследование позволило продемонстрировать возможность поступления и идущего через систему межклеточных контактов распространения наночастиц меди в тканях модельных растений, а также обуславливаемое этим глубокое повреждение ДНК. Результатом подобного механизма биологической активности наночастиц является митотоксический эффект (подавление размножения растительных клеток до вхождения в митоз), а его итоговым проявлением – фитотоксический эффект, заключающийся в подавлении роста корней Triticum aestivum и Allium cepa. Тем самым полученные результаты явились основанием для постановки вопроса о механизмах ДНК-повреждающего действия наночастиц меди, детали которого были проанализированы на модели репортерных люминесцирующих биотестов. 4.2. Исследование механизмов ДНК-повреждающего эффекта наночастиц меди в тестах на люминесцирующих штаммах Escherichia coli Исследование механизмов биологической активности наночастиц меди в отношении бактериальных клеток было начато с выяснения причин, лежащих в основе взаимодействия между ними и по предварительным данным определяемых электростатическими (кулоновскими) силами. 76 Так предварительное исследование поверхностного заряда / дзета-потенциала (ζ) и размерных характеристик наночастиц меди, проведенное с использованием технологии M3-PALS (переменного электрического поля с фазовым и частотным анализом рассеянного света), позволило определить ряд особенностей их существования в сформированной коллоидной системе. При этом было констатировано наличие у наночастиц Cu положительного поверхностного заряда для 90.3 % проанализированных частиц, характеризуемого величиной ζ = +15.9±8.63 мВ (рисунок 10а). А Б Рисунок 10 - Распределение поверхностного заряда (дзета-потенциала) у наночастиц меди (а) и клеток E.coli K12 TG1 (б). Однако, умеренное номинальное значение данного параметра, оказывающегося почти в два раза меньше порогового значения ±30 мВ как условия стабильности дисперсии, определяло тенденцию к объединению 77 наночастиц меди в более крупные агрегаты. В частности, использование технологии NIBS (неинвазивного обратного рассеяния), дополненное результатами атомно-силовой микроскопии, характеризовало размер 93.5 % подобных агрегатов величиной 432.0±202.7 нм с коэффициентом полидисперсности формируемых ими суспензий, равным 0.322. В свою очередь проведение аналогичного исследования клеток сенсорного штамма E.coli K12 TG1 характеризовало их как достаточно однородные объекты размером 1069.0±162.5 нм со значениями ζ = -50.0±9.35 мВ (рисунок 10б). Различия в знаке дзета-потенциала у наночастиц меди и клеток E.coli K12 TG1 представляются наиболее вероятной причиной взаимодействия между ними, документированного с использованием атомно-силовой микроскопии (рисунок 11) и заключающегося в формировании электростатического контакта агрегатов наночастиц Cu с бактериальной поверхностью. Результатом этого являлось формирование тесного физического контакта наночастиц с поверхностными мембранными структурами бактериальных клеток. Рисунок 11 - Взаимодействие наночастиц меди с поверхностью клеток E.coli K12 TG1. Обозначения: 1 – агрегаты наночастиц меди; 2 – контактирующие с ними бактериальные клетки; 3 – интактные бактериальные клетки. 78 Другим следствием подобного взаимодействия являлось изменение номинального значения дзета-потенциала бактериальных клеток, развивающееся во времени существования смесей «E.coli K12 TG1 + наночастицы Cu» (рисунок 12). Рисунок 12 - Результаты оценки дзета-потенциала клеток E. coli K12 TG1 после 0 минут (А), 30 минут (Б) и 90 минут (В) контакта с наночастицами меди. 79 Так к 30-й минуте контакта среднее значение данного параметра составило ζ = -23.7±12.2 мВ при его неизменности в контроле. Одновременно зафиксировано асимметричное распределение величины дзета-потенциала в популяции E.coli K12 TG1, оставшегося на уровне -43.1±5.78 мВ у 20.3 % исследованных бактериальных клеток и снизившегося до -19.3±7.78 мВ у прочих 79.7 % анализируемых объектов. При этом объяснением данного факта может являться вероятностный характер электростатического взаимодействия в системе «E.coli K12 TG1 + наноCu», сохраняющий определенную долю бактериальных клеток в интактном состоянии (рисунок 11). На этом фоне увеличение продолжительности контакта до 90 минут вело к дальнейшему снижению среднего значения дзета-потенциала до -10.1±24.6 мВ, что было результатом уменьшения доли клеток с его исходным значением до 6.18 %, а также предположительно появления 15.9 % являющихся объектов с микроорганизмами, ζ = +27.4±6.75 утратившими мВ, свою жизнеспособность после контакта с наночастицами меди. Тем самым полученный результат хорошо согласовывался с приведенными в разделе 3.2 результатами исследования токсичности наночастиц меди в отношении бактериальных клеток, одновременно объясняя и описывая последовательность формирующегося эффекта. С другой стороны, полученный результат, заключающийся в выявлении электростатического контакта наночастиц Cu с бактериальной поверхностью, а также следующего за этим повреждения энергетики бактериальных клеток, явился основанием для продолжения исследований тонких механизмов биоактивности наночастиц меди, потенциально являющихся универсальным механизмом их воздействия на живые, в том числе растительные системы. 1. В частности, на следующем этапе работы была проанализирована возможность развития окислительного стресса бактериальных клеток-мишеней, пусковым механизмом которого может являться интеграция электропроводящих наночастиц меди в локализованные их на цитоплазматических мембранах электроннотранспортные (дыхательные) цепи. При этом в качестве 80 бактериальных биосенсоров в данной серии экспериментов выступали репортерные люминесцирующие штаммы на основе E.coli K12 MG1655, несущие плазмиды с генными слияниями pSoxS::lux или pKatG::lux и ранее использованные для идентификации аналогичных механизмов биологической активности наночастиц TiO2 (Завильгельский Г.Б., Котова В.Ю., Манухов И.В. Наночастицы диоксида титана (TiO2) индуцируют в бактериях стрессовые реакции, фиксируемые специфическими lux-биосенсорами // Российские Нанотехнологии. 2011. Т. 6. №5–6. С. 401-405). Полученные результаты позволили констатировать, что контакт бактериальных клеток с частицами наночастицами Cu в диапазоне концентраций 0.625 – 5 мМ ведет к образованию в них активных форм кислорода (АФК), регистрируемому на основе специфически запускаемой с промоторов PsoxS и PkatG транскрипции кассеты репортерных luxCDABE-генов как реакции на супероксиданион и перекись водорода соответственно (рисунок 13 а, б). Рисунок 13 - Кратности индукции свечения (Iинд) репортерных штаммов E.coli K12 MG1655 с плазмидами pSoxS::lux (а), pKatG::lux (б) и pRecA::lux (в) при контакте с наночастицами (1), ионами (2) и микрочастицами (3) меди. 81 При этом более выраженная максимальная индукция генноинженерной конструкции soxS::lux (Iинд = 9.23±0.46) по сравнению с katG::lux (Iинд = 6.49±0.33) может быть интерпретирована как свидетельство первичности образования супероксиданиона, потенциально возникающего при переносе электрона через электропроводящую наночастицу меди на молекулярный кислород и в дальнейшем спонтанно дисмутирующего в перекись водорода. С другой стороны, представленные на рисунке 13 данные свидетельствуют о том, что способность к индукции окислительного стресса у микроорганизмов не является исключительным свойством наночастицам меди, но присуща и иным сравниваемым соединениям данного металла. При этом значимые особенности подобной активности заключались в том, что в присутствии ионов Cu2+ транскрипция генов-репортеров с промоторов PsoxS и PkatG запускалась при более низких (0.02 – 1.25 мМ), а в присутствии микрочастиц меди при более высоких (5 – 10 мМ) концентрациях этого металла, а достигаемые максимальные кратности индукции свечения оказывались в 1.5-3 меньше таковых при использовании наночастиц Cu. В завершение данной серии экспериментов с использованием репортерного штамма E.coli pRecA::lux, отвечающего индукцией свечения при развитии SOSответа, установлено подобное следствие воздействия наночастиц меди на бактериальные клетки-мишени как результата повреждения их ДНК образующимися в процессе окислительного стресса активными формами кислорода (рисунок 13в). При этом развитие свечения регистрировалось в широком диапазоне концентраций (1.25 – 5 мМ) и характеризовалось наиболее высокими кратностями индукции (Iинд = 15.54±0.77), что позволяло рассматривать ДНК-повреждающее действие в качестве ведущего механизма токсичности наночастиц Cu. В свою очередь сопоставимый уровень индукции репортерной конструкции recA::lux при воздействии ионов Cu2+ при меньшей выраженности окислительного стресса мог определяться развитием реакций Фентона и ГабераВейса, эффективно генерирующих гидроксил-радикалы с выраженной способностью к глубокому повреждению генетического аппарата клеток- 82 мишеней. С другой стороны, при контакте с микрочастицами меди подобная реакция была минимальной, что определило слабую биоактивность данных соединений. Таким образом, совокупность экспериментальных результатов, полученных с использованием растительных и бактериальных биотестов, позволили сформулировать единую концепцию воздействия наночастиц меди на живые системы и обусловленные этим их адаптивные реакции, схематически представленную на рисунке 14. Рисунок 14 - Предполагаемый механизм адаптации растений к действию наночастиц меди (пояснения в тексте). 83 В качестве «пускового» момента подобной биоактивности полученные данные позволяют считать электростатическое взаимодействие положительно заряженных металлических наночастиц с поверхностью клеток как про-, так и эукариот, в аналогичных условиях характеризующихся различным по значению, но идентичным по знаку отрицательным дзета-потенциалом. При этом если в случае микроорганизмов структурные препятствия для подобного взаимодействия отсутствуют, то у растений наиболее вероятным местом локализации данного контакта является ризодерма (эпиблема), покрывающая молодые коневые окончания и активно поглощающая элементы минерального питания, в то время как в экзодерме и коре этот процесс существенно затруднен или даже невозможен. В свою очередь дальнейшее распространение наночастиц металлов внутри корня растений происходит по системе межклеточных контактов, где лимитирующим фактором является диаметр плазмодесм, «разрешающий» миграцию наночастиц только сопоставимого или меньшего линейного размера. Дальнейшая реализация биоактивности наночастиц меди предполагает их интеграцию в действующие в клетках электронно-транспортные цепи. В случае микроорганизмов подобный феномен потенциально возникает практически сразу после взаимодействия наночастиц с бактериальной поверхностью, в периплазматическом пространстве под которой находятся многочисленные переносчики электронов, а в ниже расположенной цитоплазматической мембране – ключевые белки электронно-транспортных цепей. В случае же растительных клеток условием для подобной интеграции является взаимодействие наночастиц с поверхностью митохондрий. При этом в обоих обсуждаемых случаях положительный заряд наночастиц меди в совокупности с их высокими элекронно-проводящими свойствами обуславливают «ошибочный» перенос электронов через тело наночастиц на молекулярный кислород с возникновением супероксид-аниона, в дальнейшем спонтанно дисмутирующего в перекись водорода и далее превращающегося в гидроксил-анион. 84 Появление в клетке названных активных форм кислорода (АФК) обуславливает индукцию соответствующих систем антиоксидантной защиты, в случае недостаточности которых АФК начинают окислять внутриклеточные биополимеры, в том числе ДНК. При этом в случае микроорганизмов подобный механизм является непосредственной причиной гибели бактериальных клеток, а в случае растений, наряду с клеточной гибелью, приводит к прекращению деления сублетально поврежденных растительных клеток, что экспериментально регистрируется как митотоксический эффект наночастиц меди. Наконец его итоговым проявлением является фитотоксический эффект, заключающийся в прекращении роста корня в длину, адаптивном образовании боковых корней, а в своем крайнем проявлении – в гибели всего растительного организма. Тем самым, полученные результаты заставляют рассматривать зарегистрированную биологическую активность наночастиц меди как проявление универсальных свойств, реализуемых в отношении объектов разного уровня организации (Blinova I., Ivask A., Heinlaan M., Mortimer M., Kahru A. Ecotoxicity of nanoparticles of CuO and ZnO in natural water // Environ. Pollut. 2010. Vol. 158 (1). P. 41-47; Heinlaan M., Ivask A., Blinova I., Dubourguier H.C., Kahru A. Toxicity of nanosized and bulk ZnO, CuO and TiO2 to bacteria Vibrio fischeri and crustaceans Daphnia magna and Thamnocephalus platyurus // Chemosphere. 2008. Vol. 71 (7). P. 1308-1316). При этом охарактеризованная природа биоактивности наночастиц Cu оказывается во многом сходной с таковой у наночастиц TiO2 и ZnO (Завильгельский Г.Б., Котова В.Ю., Манухов И.В. Наночастицы диоксида титана (TiO2) индуцируют в бактериях стрессовые реакции, фиксируемые специфическими lux-биосенсорами // Российские Нанотехнологии. 2011. Т. 6. №5–6. С. 401-405; Kumar A., Pandey A.K., Singh S.S., Shanker R., Dhawan A. Engineered ZnO and TiO2 nanoparticles induce oxidative stress and DNA damage leading to reduced viability of Escherichia coli // Free Radical Biology and Medicine. 2011.Vol. 51 (10). P. 1872-1881), что свидетельствует в пользу существенной близости механизмов действия электропроводящих наночастиц металлов и их оксидов в отношении живых систем. Одновременно полученные результаты 85 частично позволяют частично объяснить отсутствие токсических свойств у наночастиц железа, вследствие выраженного окисления их поверхности до Fe3O4, α-Fe2O3 и γ-Fe2O3, утративших электропроводящие свойства и неспособных реализовывать описанный выше для наночастиц меди механизм биологической активности. В этих условиях наночастицы железа начинают функционировать в качестве элементов минерального питания, снабжая растительные организмы биодоступным Fe, что в ряде случаев сопровождается уже не рост- ингибирующим, но, напротив, рост-стимулирующим эффектом, исследование которого явилось предметом завершающего этапа настоящей работы. 86 ГЛАВА 5. АДАПТИВНЫЕ РЕАКЦИИ TRITICUM AESTIVUM К ПРИСУТСТВИЮ В СРЕДЕ КУЛЬТИВИРОВАНИЯ НАНОЧАСТИЦ ЖЕЛЕЗА Задачей завершающего этапа настоящего исследования явился сравнительный анализ биологической активности наночастиц железа с ионной и микроструктурированной формами этого металла в тесте прорастания семян пшеницы Triticum aestivum, рекомендуемом действующим национальным нормативом (Методические указания 1.2.2635-10 // Медико – биологическая оценка безопасности наноматериалов. – М.: Федеральный центр гигиены и эпидемиологии Роспотребнадзора. 2010, 123 с.) для аттестации биологической активности наночастиц и наноматериалов, с акцентом на выявление адаптивных реакций взаимодействующих с ними растительных организмов. Проведение соответствующих экспериментов с внесением различных форм железа в деминерализованный песочный субстрат позволило констатировать, что в отличие фитотоксического эффекта ионной формы, нано- и микрочастицы данного металла не изменяли частоту прорастания семян Triticum aestivum, а наночастицы оказывали на развитие формирующихся растений умеренно выраженное стимулирующее воздействие, нелинейно зависящее от концентрации данного соединения в среде культивирования. Так инкубация семян Triticum aestivum в контакте с ионами железа в концентрации 100 ммоль/л полностью подавляла их прорастание, а в концентрации 25 ммоль/л снижала частоту прорастания до 13,3±6,8% против 98,3±2,6% в контроле (P<0,001). При этом подобное воздействие полностью блокировало развитие корневой системы, одновременно почти десятикратно сокращая скорость роста побега: до 0,04 мм/ч против 0,47 ± 0,03 мм/ч в контроле (P<0,001). Дальнейшее уменьшение концентрации FeSO4 в среде культивирования до 6,25 ммоль/л сопровождалось увеличением частоты прорастания семян пшеницы до контрольных значений, но продолжало негативно сказываться на скорости роста побегов и корней образующихся проростков (таблица 7). 87 Регистрируемыми проявлениями в данном случае являлись двукратное снижение скорости роста побега (0,21 ± 0,02 мм/ч; P<0,05), а также значительным Таблица 7 - Скорость роста Triticum aestivum при проращивании в присутствии ионов, нано- и микрочастиц железа (воздействие 168 ч) Концентрация Исследуемые вещества веществ в субстрате, FeSО4×7H2О наночастицы Fe микрочастицы Fe ммоль/л Скорость роста побегов (мм/ч) Контроль 0,47 ± 0,03 100 0 0,47± 0,03 0,48 ± 0,04 25 0,04 ± 0,01*** 0,52 ± 0,04* 0,51 ± 0,08 6,25 0,21 ± 0,02* 0,51 ± 0,03* 0,52 ± 0,06 1,56 0,45 ± 0,02 0,48 ± 0,04 0,47 ± 0,07 0,39 0,47 ± 0,03 0,48 ± 0,03 0,45 ± 0,05 Скорость роста корней (мм/ч) Контроль 1,50 ± 0,09 100 0 1,22±0,10 1,60 ± 0,15 25 0 1,77± 0,12* 1,64 ± 0,08 6,25 0,023± 0,002*** 1,80±0,11** 1,67 ± 0,05 1,56 1,09 ± 0,15 1,72±0,10* 1,64 ± 0,14 0,39 1,36 ± 0,14 1,63±0,12 1,70 ± 0,09 Обозначения: * - P<0,05; ** - P<0,01; *** - P<0,001 подавлением скорости роста корневой системы (до 0,023 мм/ч против 1,5 мм/ч в контроле; P<0,001). Дополнительной адаптивной реакцией Triticum aestivum на присутствие в среде культивирования названной субтоксической концентрации ионов железа являлось увеличение количества придаточных корней, призванных 88 компенсировать функцию первичного корня, однако их рост в присутствии 6,25 ммоль/л FeSO4 также существенно угнетался (таблица 8). Таблица 8 - Длина первичного корня, количество, длина придаточных корней Triticum aestivum проращивании в присутствии ионов, нано- и микрочастиц железа Концентрация Длина первичного Количество Длина придаточных ммоль/л корня, мм придаточных корней, мм корней, шт Контроль 98,96 ± 5,54 2,13 ± 0,09 76,5 ± 4,12 FeSО4×7H2О 100 0 0 0 25 0 0 0 6,25 4,10 ± 0,07*** 2,84±0,11** 1,70±0,07 1,56 63,41 ± 7,05 3,40 ± 0,17 22,70 ± 4,55 0,39 83,33 ± 5,76 2,40 ± 0,09 55,01 ± 3,49 наночастицы Fe 100 83,7 ± 8,68 2,30 ± 0,18 74,6 ± 4,65 25 114,7 ± 8,29* 2,20 ± 0,17 88,2 ± 4,79* 6,25 99,5 ± 8,74 2,2 ± 0,18 89,4 ± 7,45** 1,56 97,5± 8,52 2,2 ± 0,15 87,1 ± 4,75 0,39 109,3± 7,41 2,33 ± 0,14 74,3 ± 3,88 микрочастицы Fe 100 92,68 ± 8,26 2,23 ± 0,13 79,2 ± 4,69 25 98,57 ± 7,99 2,14 ± 0,13 84,4 ± 4,97 6,25 92,80 ± 7,00 2,26 ± 0,08 81,8 ± 4.29 1,56 97,35 ± 7,51 2,30 ± 0,13 85,1 ± 4,85 0,39 96,93 ± 8,88 2,13 ± 0,16 89,0 ± 5,42 Обозначения: * - P<0,05; ** - P<0,01; *** - P<0,001 89 Дальнейшее уменьшение присутствия ионов железа в среде культивирования до 1,56 ммоль/л и ниже уже не оказывало достоверного влияния на частоту прорастания и развитие образующихся проростков Triticum aestivum. На этом фоне нано- и микрочастицы железа не изменяли частоту прорастания семян Triticum aestivum, а наночастицы оказывали на их развитие стимулирующее воздействие, которое, однако, нелинейно зависело от концентрации данного соединения в среде культивирования (таблица 7). Так значимое увеличение скорости роста побегов на 8,5-10,6 % выше контрольных значений (P<0,05) было зафиксировано в диапазоне воздействующих концентраций наночастиц железа от 25 до 6,25 ммоль/л. В свою очередь в максимальной использованной концентрации наночастицы железа всетаки несколько снижали скорость роста корневой системы Triticum aestivum, а ее наиболее выраженное увеличение скорости ее роста (до 1,80±0,11 мм/ч против 1,50 ± 0,09 мм/ч в контроле; P<0,01) фиксировалось при концентрации наночастиц железа в среде культивирования 6,25 ммоль/л с сохранением подобного стимулирующего эффекта на статистически значимом уровне в диапазоне концентраций от 25 ммоль/л до 1,56 ммоль/л. При этом отдельным аспектом полученного результата являлось наиболее выраженная стимуляция роста придаточных корней, по своей длине становящихся сопоставимыми с длиной первичного корня (таблица 8). Присутствие наночастиц железа в среде культивирования сопровождалось интенсивным поступлением и существенным накоплением данного металла в тканях Triticum aestivum, по своим абсолютным значениям превышающим аналогичные значения в тканях контрольных растений, выращенных на деминерализованном песочном субстрате (таблица 9). При этом значимым результатом являлось преимущественное накопление железа в корневой системе модельных растений, в идентичных образцах в 3,47 – 1,15 раз превышающее аналогичные значения в тканях зеленых побегов, тем самым подтверждая представления о корневой системе растений как основной «мишени» для воздействия металлических наночастиц. 90 Второй важный аспект полученного результата определялся дозозависимым характером накопления использованного металла, вновь нелинейно зависящим от концентрации наночастиц железа в среде культивирования. Таблица 9 - Количественное содержание железа в проростках Triticum aestivum (в пересчете на сухую биомассу, мкг/г), инкубированных в присутствии различных концентраций наночастиц Fe Исследуемые Концентрация наночастиц железа части Triticum в среде культивирования, мкг/мл aestivum Побеги Корни Контроль 5585 1396,2 349,1 87,2 21,8 52,24± 71,45± 39,95± 27,31± 25,9± 9,22 11,49** 15,72** 5,79* 4,80 4,00 ±3,04 181,53± 102,99± 75,09± 35,79± 25,18± 4,48 16,52*** 7,87** 5,54** ±1,48 39,93*** 22,66*** Обозначения: * - P<0,05; ** - P<0,01; *** - P<0,001 Так преимущественное накопление железа регистрировалось в корневой системе модельных растений с максимумом 181,53±39,93 мкг в 1 г сухого веса растительной биомассы при проащивании в среде с содержанием 100 ммоль/л (=5585 мкг/мл) соответсвующих наночастиц, что более чем трехкратно превышало содержание железа в тканях побега. Однако, эффективность извлечения железа из среды культивирования в данном случае оказывалась минимальной, прогрессивно увеличиваясь с 1,94 % до 19,9 % при уменьшении воздействующих концентраций наночастиц со 100 до 0,39 ммоль/л (=21,8 мкг/мл). Сказанное свидетельствует о существовании определенных ограничений поступления и распространения наночастиц железа в тканях модельных растений, предположительно вовлеченных в систему их адаптивных реакций при проращивании в условиях интенсивного контакта с наночастицами минерального происхождения. 91 Таким образом, результаты проведенного исследования свидетельствуют о выраженных различиях адаптивной реакции растений на воздействие железа в ионной и металлической (нано- и микро-) формах, продемонстрированных на модели проращивания семян Triticum aestivum. При этом наночастицы железа не проявляют выраженного токсического действия, но напротив, в определенных диапазонах концентраций стимулируют рост и развитие проростков Triticum aestivum, что сопровождается накоплением в тканях модельных растений значительных количеств воздействующего металла. Подобный результат определяет перспективу использования наночастиц железа при предпосевной обработке семян, а также в системе выращивании кормовых и сельскохозяйственных растений методами аэропоники или гидропоники, ожидаемым следствием чего явится не только повышение выхода биомассы, но и ее обогащение биологически доступным железом, востребованным при коррекции микронутриентной обеспеченности человека и животных. 92 ЗАКЛЮЧЕНИЕ По различным оценкам история жизни на нашей планете включает около 2 млрд. лет и на протяжении всего этого периода живые системы вынуждены непрерывно приспосабливаться к условиям окружающей среды. Столь долгий срок наложил свой отпечаток на организмы, сформировав их как динамичные, хорошо адаптированные к внешним условиям системы органов и тканей. Не стали исключениями и высшие растения. В настоящей работе мы попытались осветить только один из аспектов приспособления растений к внешней среде - связанный с особенностями их минерального питания. Важность данного фактора в жизни растений неоднократно подчеркивалось многими учеными (Трапезников В.К, Иванов И.И., Тельвинская Н.Г. Локальное питание растений. Уфа: Гилем, 1999. 260 с. и т.д.). В результате наукой накоплен значительный багаж знаний, касающийся изменений в архитектонике и морфологии растений, находящихся в условиях дефицита или избытка компонентов минерального питания. Сегодня, хорошо известны оптимальные, для растительных организмов параметры концентраций химических элементов, что позволило человеку широко использовать технологии искусственных питательных сред. Основной целью представленной работы стало продолжение исследований в данном направлении, а именно - изучение приспособления высших растений к условиям присутствия в среде различных источников эссенциальных элементов, в т.ч. вещества в наноформе. Нельзя сказать, что высшие растения некогда в своей эволюции «не сталкивались» с наличием высокодисперсных веществ. Напротив, вся история этих организмов неразрывно связана со взаимодействием с микро- и наночастицами минерального происхождения. Выше мы уже приводили данные такого тесного взаимодействия, они действительно впечатляют. В атмосферном воздухе содержится до 1 млн минеральных частиц размером от 0,001 до 1000 мкм., в пресной и морской воде их концентрация может достигать 5-6 млн/дм3 (Yushkin N.P. Mineralis mundi et biosphere: mineralis organiz-mobioz, biomineral interaction coevolution // Acta IV International seminar «Mineralogy et vitam: Origin 93 de biosphere et co-evolution of mineralis et biologicum mundos biomineral-ogiya». Syrtyvkar, 2007. Р. 5-7). Безусловно, эволюционно высшие растения адаптировались к таким условиям. Однако, по мере развития нанотехнологии в окружающую среду поступает все большее количество ранее неизвестных наноматериалов, перечень которых уже сейчас превышает 4 тыс. наименований. Сказанное определяет актуальность детального изучения механизмов приспособления высших растений, как неотъемлемой части экосистемы, к техногенным высокодисперсным веществам. Необходимость проведенных ботанических исследований продиктована и интересами сельского хозяйства. В настоящее время наночастицы металлов получают все большее распространение в животноводстве в качестве источников микроэлементов (Мирошникова Е.П., Аринжанов А.Е., Глущенко Н.Н., Василевская С.П. Обмен химических элементов в организме карпа при использовании наночастиц кобальта и железа в корме // Вестник Оренбургского гос. ун-та. 2012. №6. С. 55-57; Мирошникова Е.П., Аринжанов А.Е., Килякова Ю.В. Изменение гематологических параметров карпа под влиянием наночастиц металлов // Достижения науки и техники АПК. 2013. №5. С. 55-57). Эти формы, при определенных характеристиках, оказались менее токсичными чем традиционные соли минеральных солей и относительно более дешевыми, чем органические формы (Глущенко Н.Н. Физико-химические закономерности биологического действия высокодисперсных порошков металлов: автореф. дис. дра биол. наук. М., 1989). Кроме того, наночастицы все чаще используются и в растениеводстве, в частности - как источники химических элементов при предпосевной обработке семян (Крылов С.В., Паничкин Л.А., Захарин А.А. и др. Способ оценки метода протравливания семян // Патент РФ №2015631 от 15.07.1994 г; Черняев Н.Г. Изучение влияния препаратов и способов обработки семян на перезимовку озимых зерновых культур // Известие ТСХА. 1997. Вып.2. С.23-28; Райкова А.П. Предпосевная обработка семян // Материалы международной научно- 94 практической конференции «Льняной комплекс россиии, проблемы и перспективы. Вологда, 2001; Райкова А.П., Паничкин Л.А., Райкова Н.Н. Использование ультрадисперсных порошков металлов для предпосевной обработки семян // Доклады ТСХА. 2004. Вып. 276. С. 44-48; Райкова А.П., Паничкин Л.А., Райкова Н.Н. Исследования влияния ультрадисперсных порошков металлов, полученные различными способами на рост и развитие растений // Материалы междунар. науч.-техн. конф. «Нанотехнологии и информационные технологии – технологии 21 века». М., 2006. С. 118-123). Однако, нельзя не помнить и о других уникальных свойствах наночастиц, в спектре которых присутствуют и токсические эффекты. Следствием этого в последние годы стало бурное развитие нанотоксикологии. Эта наука появилась после первых публикаций, датируемых 2002-2006 годами, и посвященных токсическому действию наночастиц на различные организмы. Несмотря на относительную «молодость» эта наука за десять лет своего существования накопила большой багаж знаний о токсических аспектах воздействия наночастиц. Необходимость формирования этой дисциплины продиктована объективными причинами, в числе которых невозможность оценки токсичности наночастиц по сравнению с аналогами в макродиспесной форме или в виде сплошных фаз, так как токсикологические свойства наноматериалов являются результатом не только их химического состава, но и разнообразия других физико-химических особенностей. Имеющиеся токсикологические методологии основаны на определении токсичности вещества относительно массовой концентрации, что неприемлемо для наноматериалов, для которых одним из основных определяющих свойств является величина площади поверхности или число наночастиц. Работая с наноматериалами, исследователи вынуждены непрерывно формировать новые базы данных, обеспечивающие выявление новых индикаторов нанотоксичности, которые должны обязательно учитывать вклад таких характеристик, как: размер, форма, состав, химическая реактивность составляющих их частиц и т.д. Однако до сих пор методы выявления, идентификации и количественного определения наноматериалов в объектах 95 окружающей среды, пищевых продуктах и биосредах, которые могли бы достоверно отличить их от химических аналогов в макродисперсной форме, разработаны недостаточно. Исходя из вышесказанного, нами были запланированы и проведены представляемые Вашему вниманию исследования по оценке особенностей роста, развития и приспособление высших растений к условиям среды, значимым компонентом которой являются наночастицы металлов. При планировании экспериментов в качестве тест-объектов наночастиц нами были выбраны две широко использованные в подобных ботанических исследованиях биологические модели: Triticum aestivum и Allium сера L. В свою очередь в качестве инструмента воздействия были выбраны наночастицы (80-100 нм), микрочастицами (10-40 мкм) и солями этих металлов. сравниваемые с По окончанию исследований можно констатировать, что необходимые требования по точности измерений и чистоте экспериментов, на наш взгляд, были выполнены. Во-первых, выполнение основных экспериментов по оценке особенностей роста и развития растений, а также реакций микроорганизмов было проведено с учетом методических указаний МУ 1.2. 2635-10 «Медико-биологическая оценка безопасности наноматериалов». М.: Федеральный центр гигиены и эпидемиологии Роспотребнадзора, 2010 и МУ 1.2.2634- 10 «Микробиологическая и молекулярно-генетическая представителей оценка микробиоценоза». М.: воздействия Федеральный наноматериалов центр гигиены на и эпидемиологии Роспотребнадзора, 2010. Во-вторых, основные измерения и испытания были проведены на базе технически компетентных экспериментальных площадок: Испытательного Центра ВНИИ мясного скотоводства (аккредитация Госстандарта России Росс. RU 000121 ПФ59) и Центров коллективного использования «Институт микр- и нанотехнологий» (Оренбург) и «Симбиоз» (Саратов) на поверенном оборудовании. . В-третьих, использованные наночастицы содержали химически чистые металлы, с содержанием примесей от 0,05 до 0,001 %. Предварительная оценка 96 основных характеристик данных наночастиц была проведена с использованием широкого спектра методов в условиях испытательного центра Института энергетических проблем РАН. В процессе исследований была установлена неоднозначная реакция растений на присутствие оцениваемых веществ в среде, что отмечалось уже на этапе оценки всхожести зерен пшеницы и прорастания луковиц. Инкубация семян Triticum aestivum в контакте с ионами железа в концентрации 100 ммоль/л вела к гибели зародышей, а в концентрации 25 ммоль/л снижала частоту прорастания на 85,0 % (P<0,001). Между тем снижение концентрации ионов сопровождалось восстановлением всхожести, на фоне которого выраженные изменения были констатированы при анализе размерных характеристик побегов и корней проростков. Так концентрация ионов железа 25 ммоль/л блокировала развитие корневой системы, одновременно почти десятикратно сокращая длину побега. В свою очередь концентрация ионов железа 6,25 ммоль/л снижала длину корней до 16,0±0,1 мм против 221,4±21,3 мм. Рассчитанные на данной основе значения EC50, соответствующие подавлению результативного параметра на 50% от контрольных значений и тем самым характеризующие верхний предел толерантности Triticum aestivum к ионам железа, свидетельствовали об увеличении выраженности токсического эффекта в ряду «всхожесть семян → длина побега → длина корня». С другой стороны, в ходе исследований нами не было выявлено достовернго подавления роста пшеницы мягкой по мере роста концентраций нано- или микрочастиц железа. Напротив, следствием дополнительного введения нанометаллов, стало увеличение основных ростовых параметров. Подобная реакция растений представляется вполне закономерной. Так, по данным ряда исследований отсутствие в среде минеральных веществ (в нашем случае контроль) сопряжено со снижением роста растений. Важным в этой связи является компенсаторная реакция растений пшеницы мягкой на условия минерального питания, сводящаяся к изменению относительной скорости побегов и корневой системы. 97 Выбор контрольной модели минерального питания как ноль-вариант позволил нам сформировать гомологический ряд веществ-источников микроэлементов. Оказалось, что в нетоксических дозах выбранные источники оказались не однозначными по данному признаку. При этом в наибольшей степени ростостимулирующими эффектами отличались именно наночастицы железа как важнейшего биогенного элемента, играющего ключевую роль во многих метаболических и энергетических процессах про- и эукариот. По нашему мнению наночастицы данного металла могут функционировать в качестве «медленного генератора» ионов железа, на физиологическом уровне усваиваемых растениями в динамике их роста и развития в противоположность разовому поступлению таких ионов в виде солей, оказывающих неблагоприятный (токсический) эффект. Иная ситуация зафиксирована нами при сравнительном исследовании нанои микрочастиц меди с эффектами ионной формы этого металла. Проведенные исследования свидетельствовало о досточно низкой толерантности Triticum aestivum к подобному воздействию. В частности, анализ морфологии проростков Triticum aestivum, инкубированных в присутствии наночастиц Cu, позволил констатировать статистически значимое изменение ряда биометрических параметров, преимущественно относящихся к корневой системе данного растения. В свою очередь наночастицы меди хотя достоверно и не изменяли частоту прорастания семян, но в максимальной использованной концентрации 100 ммоль/л негативно влияли на длину побегов, а наиболее выражено подавляли формирование корневой системы. Помимо этого результатом воздействия наночастиц меди на Triticum aestivum являлось уменьшение массы, увеличение диаметра образующихся корней, а также их необычное ветвление по типу «куриной лапки». На этом фоне еще меньшая толерантность была зарегистрирована к ионной форме меди, выражено подавляющей частоту всхожести семян Triticum aestivum (EC50=4 ммоль/л), а также достоверно уменьшающей длину формирующихся побегов (EC50=3,5 ммоль/л) и корней (EC50=0,2 ммоль/л), что в 2-4-16 раз 98 превышало аналогичные значения для ионов железа. В свою очередь для микрочастиц Cu аналогичная величина была выявлена только для корневой системы EC50=6,7 ммоль/л. Как известно протравливание ионами Сu2+ уже с начала XX века стало распространенным методом предпосевной защиты растений. В, соответствии, с чем результаты настоящего исследования свидетельствуют о возможности реализации аналогичного подхода и с использованием наночастиц меди. В этой связи органичным продолжением работ стало исследование пределов толерантности рекомбинантного люминесцирующего штамма Escherichia coli к оцениваемым факторам, что дополнительно было обусловлено использованием этого организма в качестве модели при изучении взаимодействий в системе «растение-наночастицы». В целом результаты этих экспериментов были сходными с описанными выше для растений. Так контакт E. coli с возрастающими концентрациями ионов меди в диапазоне от 100 до 0,78 ммоль/л вел к развивающемуся в первые 15-60 минут выраженному токсическому эффекту - полному подавлению бактериального свечения. В свою очередь для формирования аналогичного эффекта ионов меди в концентрации 0,39 ммоль/л требовалось уже 170 мин, а еще более низкие дозировки обуславливали лишь относительную ингибицию бактериальной биолюминесценции до 10-70 % от контрольных значений. Наночастицы меди также обуславливали ингибирование бактериальной биолюминесценции, количественные характеристики которой, однако, имели ряд существенных отличий. Первым из них было замедленное формирование токсического эффекта, развивающегося только со 120-й минуты контакта. Второй особенностью стало то, что для достижения подобного эффекта требовались значительно более высокие концентрации наночастиц меди - от 12,5 до 100 ммоль/л. Таким образом, результаты исследований позволили получить развернутые представления о значениях верхнего предела толерантности модельных растений и бактерий к препаратам железа и меди, закономерно снижающейся в ряду 99 «микрочастицы → наночастицы → ионы» и нарастающей от меди к железу. При этом детали подобного эффекта заключались в идентификации корневой системы растений как основной «мишени» для воздействия металлических наночастиц, формирующей основу для продолжения исследований в заданном направлении для выявления механизмов регистрируемого фитотоксического эффекта. Важным результатом стало и то, что микроорганизмы оказывались более чувствительными к действию исследуемых соединений металлов чем растения, однако, существенный параллелелизм результатов исследования биологической активности ионов, нано- и микрочастиц в растительном биотесте и тесте ингибирования бактериальной биолюминесценции определил возможность последующего использования данных о тонких механизмах токсичности названных соединений, получаемых с использованием бактериальных моделей, и для объяснения характера их действия в отношении высших растений. Как было показано выше, адаптивные реакции Triticum aestivum к присутствию в среде культивирования наночастиц меди, в первую очередь проявлялось в изменениях корневой системы растений. Указанное обстоятельство определило интерес к углубленному изучению механизмов и последствий взаимодействий в системе «корень-наночастицы», реализованному с использованием микроскопических и генетических методов анализа. Электронно-микроскопическое исследование контрольных образцов апикальной части корней Triticum aestivum выявляли ряд ультраструктурных изменений, затрагивающих характер организации клеточных стенок. При этом в них обнаруживались многочисленные плазмодесмы, визуализируемые как электронно-плотные полосы, пересекающих клеточные стенки на расстоянии 0,52 мкм друг от друга. В использованном экспериментальном контексте (в отсутствие дополнительного контрастирования) формирование электронной плотности на участках плазмодесм могло быть следствием прохождения через них наночастиц, оставляющих за собой «следы» из дислоцировавших с поверхности кластеров меди. Дополнительными аргументами в пользу данного предположения являются 100 сопоставимость диаметра плазмодесм (до 70 нм) и использованных наночастиц, а также регистрируемая на некоторых электроннограммах ассоциированность последних с местами открытия плазмодесм в цитозоль растительных клеток. Кроме того, важной особенностью оказывалось более чем двукратное (684±120 нм, P<0,01) утолщение клеточных стенок в корнях, выросших в присутствии наночастиц меди. Данный факт, с одной стороны, может рассматриваться как одно из проявлений реакции тканей растения на проникновение и распространение наночастиц меди, а с другой – объяснять природу утолщения корней пшеницы при подобном воздействии по сравнению с соответствующим контролем. Другим важным результатом электронно-микроскопического исследования являлась визуализация в цитоплазме клеток опытных образцов корня Triticum aestivum многочисленных электронно-плотных образований округлой формы, по своим характеристикам соответствующих параметрам использованных наночастиц. Сказанное свидетельствует в пользу возможности поступления меди в ткани растений в исходной наноструктурированной форме, где предполагаемой движущей силой данного процесса является градиент концентраций наночастиц в системе «среда культивирования – ткань корня», а механизмом распространения – межклеточный перенос вглубь корня через систему плазмодесм. В продолжение анализа распределения наночастиц меди в тканях модельных растений количественное содержание этого элемента с пересчетом на сухую биомассу Triticum aestivum было исследовано методом атомно- адсорбционной спектроскопии. Исследование непосредственных механизмов снижения адаптационной способности растений к среде содержащей наночастицы меди было проведено в модифицированном варианте Allium-теста. При этом проведенный анализ контрольных образцов позволил выявить типичную для меристематических тканей корня лука картину, характеризующуюся присутствием значительного количества клеток, находящихся на различных стадиях митоза. 101 В свою очередь микроскопическое исследование корней, пророщенных в контакте с наночастицами пролиферативной свидетельствовало активности со о стремящимися полном подавлении нулю значениями к митотического индекса, сохраняющем клетки в интерфазном состоянии и делающем технически невозможным регистрацию митоз-модифицирующих или мутагенных эффектов. Дополнительным проявлением подобного митотоксического эффекта наночастиц меди являлись деструкция и конденсация хроматина, что соответствовало процессу пикнотического разрушения ядра и являлось одним из признаков гибели растительных клеток. С целью выяснения природы регистрируемых явлений в отдельной серии экспериментов действие наночастиц меди было оценено в оригинальном варианте Allium-теста, подразумевающем контакт их суспензии с предварительно пророщенной корневой системой лука. При этом электрофоретическое разделение ДНК, выделенной из концевых отрезков корней после 3-часовой инкубации в среде с наночастицами меди, позволило констатировать существенную и дозозависимую деградацию данного биополимера. В качестве вероятной причины происходящих событий было выдвинуто предположение о взаимодействии наночастиц меди с внутриклеточными цепями переноса электронов, для проверки которого была проведена серия экспериментов с клетками Escherichia coli, являющимися близкими аналогами митохондрий эукариот. При этом в качестве объектов иследования в данной серии экспериментов выступали репортерные люминесцирующие штаммы на основе E.coli K12 MG1655, несущие плазмиды с генными слияниями pSoxS::lux, pKatG::lux или recA::lux, реагирующие развитием свечения при возникновении окислительного стресса или повреждения ДНК, соответственно. Полученные результаты позволили констатировать, что контакт бактериальных клеток с наночастицами меди вел к образованию в них активных форм кислорода, регистрируемому на основе специфически запускаемой с промоторов PsoxS и PkatG транскрипции кассеты репортерных luxCDABE-генов как реакции на супероксиданион и перекись водорода соответственно. При этом 102 более выраженная максимальная индукция генноинженерной конструкции soxS::lux по сравнению свидетельство с первичности katG::lux была образования интерпретирована супероксиданиона, нами как потенциально возникающего при переносе электрона через электропроводящую наночастицу меди на молекулярный кислород и в дальнейшем спонтанно дисмутирующего в перекись водорода. В завершение данной серии экспериментов с использованием репортерного штамма E.coli pRecA::lux, отвечающего индукцией свечения при развитии SOSответа, установлено подобное следствие воздействия наночастиц меди на бактериальные клетки-мишени как результата повреждения их ДНК образующимися в процессе окислительного стресса активными формами кислорода. При этом развитие свечения регистрировалось в широком диапазоне концентраций и характеризовалось наиболее высокими кратностями индукции, что позволяло рассматривать ДНК-повреждающее действие в качестве ведущего механизма токсичности наночастиц меди. Таким образом, полученные данные демонстрируют возможность поступления и идущего через систему межклеточных контактов распространения наночастиц меди в тканях модельных растений, а также обуславливаемое этим глубокое повреждение ДНК. С использованием бактериальных моделей показана индукция окислительного стресса в присутствии наночастиц, обуславливающих образование активных форм кислорода, которые, в свою очередь, повреждают ДНК. В свою очередь, низкая адаптивная способность растений к условиям среды культивирования со значительным содержаниями наночастиц меди обусловлена повреждением (фрагментацией) ДНК растительных клеток, следствием чего является митотоксический эффект (подавление размножения клеток до вхождения в митоз), а его итоговым проявлением – фитотоксический эффект, заключающийся в нарушении роста корневой системы . Анализ полученного в исследованиях экспериментального материала позволяет предложить ряд решений по оптимизации использования растительных организмов в сельском хозяйстве и биотехнологии. Во-первых, при равной или 103 сходной стоимости различных источников микроэлементов, при формировании питательных сред растений в практической гидропонике, более целесообразным может быть использование наночастиц этих металлов. Во-вторых, учитывая различную толерантность растений и микроорганизмов к действию наноматериалов, практика использования последних может быть расширена для целей дезинфекции и предпосевной обработки семян сельскохозяйственных растений. 104 ВЫВОДЫ 1. Сравнительная оценка толерантности растений (Triticum aestivum) и микроорганизмов (Escherichia coli) в тестах прорастания семян и ингибирования бактериальной биолюминесценции демонстрирует снижение способности данных организмов к сохранению жизнеспособности при культивировании в среде, содержащей медь и железо, при нарастании негативных эффектов в ряду «микрочастицы → наночастицы → ионы металлов». 2. Растения по сравнению с микроорганизмами более устойчивы к токсическому действию меди и железа, при воздействии которых выраженность фитотоксического эффекта возрастает в ряду проявлений «всхожесть семян → длина побега → длина корня». 3. Особенностью роста и развития растений (Triticum aestivum и Allium cepa) при культивировании в среде, содержащей наночастицы меди в концентрациях более 1,5 ммоль/л, является нарушение роста и развития корневой системы растений, в т.ч. уменьшение длины первичного корня; увеличение числа придаточных корней и снижение их длины. 4. Наночастицы металлов поступают и распространяются в тканях корневой системы растений через систему межклеточных контактов (плазмодесм). Присутствие в среде культивирования наночастиц меди и железа сопровождается пропорциональным накоплением данных металлов преимущественно в корневой системе растений, повышение концентраций этих веществ в побегах менее значимо. 5. Низкая адаптивная способность растений Triticum aestivum и Allium cepa к условиям среды культивирования со значительным содержаниями наночастиц меди обусловлена повреждением (фрагментацией) ДНК растительных клеток, следствием чего является митотоксический эффект (подавление размножения клеток до вхождения в митоз), а его итоговым проявлением – фитотоксический эффект, заключающийся в нарушении роста корневой системы растений 105 6. Взаимодействие наночастиц меди с бактериальными клетками (на примере Escherichia coli) ведет к развитию у них состояния окислительного стресса, результатом которого является повреждение ДНК. 7. Присутствие наночастиц железа в среде культивирования Triticum aestivum в диапазоне концентраций от 25 о 6,25 ммоль/л сопровождается развитием ростостимулирующего эффекта, выражающегося линейного роста образующихся проростков пшеницы. в увеличении скорости 106 СПИСОК ИСПОЛЬЗОВАННОЙ ЛИТЕРАТУРЫ 1. Аринжанов, А. Е. Воздействие наночастиц комплекса металлов на организм карпа / А. Е. Аринжанов, Е. П. Мирошникова, Ю. В. Килякова // Известия Оренбургского государственного аграрного университета. – 2013. – №2. – С. 113-116. 2. Арсеньева, И. П. Аттестация наночастиц металлов, используемых в качестве биологически активных препаратов / И. П. Арсеньева // Нанотехника. – 2007. – №10. – С.72 – 77. 3. Батова, Ю. В. Накопление кадмия и его распределение по органам у растений ячменя разного возраста / Ю. В. Батова, А. Ф. Титов, Н. М. Казнина, Г. Ф. Лайдинен// Труды Карельского научного центра РАН. – 2012. №2. – С. 32–37. 4. Башмаков, Д. И. Эколого-физиологические аспекты аккумуляции и распределения тяжелых металлов у высших растений/ Д. И. Башмаков, А. С. Лукаткин. – Саранск: Изд-во Мордов. Ун-та, 2009. – 236 с. 5. Богословская, О. А. Изучение безопасности введения наночастиц меди с различными физико-химическими характеристиками в организм животных / О. А. Богословская, Е. А. Сизова, В. С. Полякова, С. А. Мирошников, Н. Н. Глущенко // Вестник Оренбургского государственного университета. – 2009. – №2. – С.124127. 6. Бритиков, Е. А. Биологическая роль пролина / Е.А. Бритиков. – М.: Наука, 1975. –88 с. 7. Ванюшин, Б. Ф. Апоптоз у растений / Б. Ф. Ванюшин // Успехи биологической химии. – 2001. – Т. 41. – С. 3-38. 8. Веселкин, Д. В. Строение и микоризация корней сеянцев ели и пихты при изменении почвенного субстрата / Д. В. Веселкин // Лесоведение. – 2002. – №3. – С.12-17. 9. Веселкин, Д. В. Реакция экомикориз Pinussylvestris L. на техногенное загрязнение различных типов / Д. В. Веселкин // Сибирский экологический журнал. – 2005. – №4. – С. 753-761. 107 10. Веселкин, Д. В. Современная микология в России / Д. В. Веселкин // Первый съезд микология в России: тез. докл. – М.: 2002. – С. 86-87. 11. Веселкин, Д.В. Экологическая токсикология: учеб. пособие / Д. В. Веселкин // Екатеренбург: Урал, 2001. – С. 38-46. 12. Веселова, Т.В. Стресс у растений (Биофизический подход) / Т. В. Веселова, В. А. Веселовский, Д. С. Чернавский. – М.: РАН, 1993. – 144 с. 13. Врублевский, Э. Нанотехнология – путь в будущее или бренд для финансирования / Э.Врублевский, В. Киреев, В. Недзвецкий // Нано- и микросистемная техника. – 2007. – № 12. – С. 6–20. 14. Гаджиева, И.Х. Кросс-адаптация растений к почвенному засолению и тяжелым металлам / И. Х. Гаджиева, З. М. Алиева, П. Б. Рамазанова // Юг России: экология, развитие. – 2010. №1. – С. 26-32. 15. Глушко, А.А. Экстремальная экология (человека и природы) / А. А. Глушко // Инженерная экология. – 2010. –№2(91). – С. 4-24. 16.Глушкова, А.В. Нанотехнологии и нанотехнология – взгляд на проблему / А. В. Глушкова, А. С. Радилов, В. Р. Рембовский // Научно-исследовательский институт гигиены, профпатологии и экологии человека ФМБА, 2007. – 30 с. 16. Глушкова, А.В. Нанотехнологии и нанотоксикология – взгляд на проблему / А. В. Глушкова, А. С. Радилов, В. Р. Рембовский // Методологические проблемы изучения и оценки био- и нанотехнологий (нановолны, частицы, структуры, процессы, биообъекты) в экологии человека и гигиене окружающей среды: материалы пленума научного совета по экологии человека и гигиене окружающей среды РАМН и Минздравсоцразвития Российской Федерации / под ред. Ю.А. Рахманина. М., 2007. – 21 с. 17.Глущенко, Н.Н. Физико-химические закономерности биологического действия высокодисперсных порошков металлов : автореф. дис. д-ра биол. наук : 14.00.33 / Глущенко Наталья Николаевна. – М., 1989. – 19 с. 18. Глущенко, Н.Н. Сравнительная токсичность солей и наночастиц металлов и особенность их биологического действия / Н. Н. Глущенко, О. А. Богословская, И. П. Ольховская // междунар. науч. – практ. конф. // 108 Нанотехнологии и информационные технологии – технологии 21 века. – М., 2006. – С. 93-95. 19. Глущенко, Н.Н. Физико-химические закономерности биологического действия высокодисперсных порошков металлов / Н. Н. Глущенко, О. А. Богословская, И. П. Ольховская // Химическая физика. – 2002. –Т.21(4). – С.79-85. Глущенко, Н.Н. Биологическое действие высокодисперсных порошков металлов / Н. Н. Глущенко, И. П. Ольховская, Т. В. Плетенева и др. // Известия РАН., Сер. Биология. - 1989. – №3. – С.415-421. 20. Голохваст, К.С. Атмосферные взвеси Владивостока: Гранулометрический и вещественный анализ / К. С. Голохваст, П. А.Никифоров, П. В. Кику и др. // Экология человека. – 2013. – №1. – С. 14-19. 21.Голохваст, К.С Влияние микрочастиц минералов на работу физиологических и биологических систем / К. С. Голохваст, И. Э. Памирский, Е. Н. Бородин и др. // Фундаментальные исследования. – 2013. – № 6. – С.909-912. 21. Голохваст, К.С. Гранулометрический и минералогический анализ взвешенных частиц в атмосферном воздухе / К. С. Голохваст, Н. К. Христофорова, П. Ф. Кику и др. // Физиология и патология дыхания. – 2011. – Вып.40. – С.94-100. 22. Голохваст, К.С. Первые данные по вещественному составу атмосферных взвесей Владивостока / К. С. Голохваст, И. Ф. Чекрыжов, А. М. Паничев и др. // Известия Самарского науного центра РАН. – 2011. – Т.13. – №1(8). – С.18531857. 23. Горышина, Т.К. Экология растений / Т. К. Горышина. – М.: Высшая школа, 1979. – 368 с. 24.Грубинко, В. В. Структурные адаптации клеточных стенок водных растений к действию цинка и свинца / В. В. Грубинко, К. В. Костюк // Бiологiчний вiсник МДПУ. – 2012. – №2. – С. 58-66. 24. Демедчик, В.В. Токсичность избытка меди и толерантность к нему растений / В. В. Демедчик, А. И. Соколик, В. М. Юрин // Успехи современной биологии. – 2001. – № 5. – Т.121. – С.511-525. 109 25. Добровольский, В.В. Химия жизни / В. В. Добровольский. – М.: 1988. – 176 с. Дыкман, Л.А. Золотые наночастицы: синтез, свойства, биомедицинское применение / Л. А. Дымкан, В. А. Богатырев, С. Ю. Щеглов и др. – М.: Наука, 2008. – 319с. 26. Егоров, И.А. Высокодисперсные порошки металлов – источники микроэлементов для сельскохозяйственной птицы / И. А. Егоров, В. П. Куренева, Н. Н. Глущенко и др. // Физиолого – биохимические основы повышения продуктивности сельскохозяйственной птицы: сборник науч. тр. - Боровск, 1985. – Т.31. – С.80-88. 27. Жигач, А.Н. Получение ультрадисперсных порошков металлов, сплавов, соединений металлов методом Гена-Миллера: история, современное состояние, перспективы / А. Н. Жигач, М. Л. Кусков, И. О. Лейпунский и др. // Российские нанотехнологии. – 2012. – Т.7(№ 3-4). – С.28—37. 28. Жмудь, Е.В. Экологическая пластичность HegysarumGmtlinii (fabactae) в Горном Алтае и Хакасии / Е. В. Жмудь // Вестник Томского Государственного политехнического университета. – 2014. – №11(152). – С. 220-226. 29. Зайцев, Г.А. Особенности формирования микоризы сосны обыкновенной в условиях промышленного загрязнения / Г. А. Зайцев, Г. М. Мухаметова, Д. В. Веселкин// Вестник Оренбургского гос. ун-та. - 2009. – №6. – С.137-139. 30. Захарин, А.А. Особенности водно-солевого обмена растений при солевом стрессе / А. А. Захарин // Агрохимия. – 1990. – № 8. – С.69-79. 31. Иванова, М.И. Использование ультрадисперсных порошков металлов и биологически активных веществ для предпосевной обработки семян :автореф.дис. … канд. биол. наук : 06.01.06 / Иванов Михаил Иванович. – М.: 1997. – 19 c. 32. Иванычева, Ю.Н. Жечкова, Т.В., Полищук, С.Д. Влияние нанопорошков меди и оксида меди на активность фитогормонов в проростках вики и яровой пшеницы / Ю. Н. Иванычева, Т. В. Жечкова, С. Д. Полищук // Вестник ФГБОУ ВПО РГАТУ. – 2012. – №1(13). – С. 12-14. 110 33. Ильин, В. Б. Тяжелые металлы в системе почва – растение / В. Б. Ильин. – Новосибирск: Наука, 1991. – 150 с. 34. Ильичев, Е. Переваримость рациона и баланс питательных веществ при скармливании телятам нанопорошков кобальта и меди / Е. Ильичев, А. Назарова, С. Полищук, В. Иноземцев// Молочное мясное скотоводство. – 2011. – №5. – С. 2729. 35. Илющенко, В.Г. Классификация спонтанной генотипической клеточной адаптации / В. Г. Ильющенко // Цитология и генетика. – 2002. –Т.36. – №5. – С. 34-42. 36. Исаева, А.Ю. Изучение биологических свойств наноразмерной структуры на основе коллоидного селена invitro/ А. Ю. Исаева, С. А. Старовертов, А. А. Волков, С. В. Ларионов, С. В. Козлов // Ветеренарная патология. – 2012. – №3. – С.111-114. 37. Кайгородов, Р.В. Загрязняющие вещества пыли проезжих частей дороги и в древесной растительности придорожных полос городской зоны / Р. В. Кайгородов, М. И. Тиунова, А. В. Дружинина // Вестник Пермского университета. – 2009. – Вып.10. – С. 141-146. 38. Керимов, Ф.А. Организменный и клеточный уровни солеустойчивости двух сортов хлопчатника [133, ИНЭБР-85] / Ф. А. Керимов, В. В. Кузнецов, З. Б. Шамина // Физиология растений. – 1993. –Т.40. –№ 1. – С.128-131. 39.Кисель, В.П. Нетрадиционные природные ресурсы, инновационные технологии и продукты / В. П. Кисель // Сб. науч. тр. - М.: РАЕН, 2003. – Вып. 10. - С. 183-196. 39.Кисель, В.П. Фитотерапия, биологически активные вещества естественного происхождения / В. П. Кисель // Матер. 5 междунар. конф. РАН. Черноголовка, 2004. – С. 170-174. 40. Князева, С.Г. Морфолого-анатомические особенности хвои можжевельника обыкновенного (JuniperuscommunisL.) / С. Г. Князева // Хвойные бореальной зоны. – 2012. – №1-2. – С. 92-96. 111 41. Кобаяси, Н. Введение в нанотехнологию / Н. Кобаяси. – М.: Бином. Лаборатория знаний, 2007. – 134с. 42. Коваленко, Л.В. Активация прорастания семян ультрадисперсными порошками железа / Л. В. Коваленко, Г. Э. Фолманис // Достижения науки и техники АПК. – 2001. – №9. – С. 7-8. 43. Коваленко, Л.В. Биологически активные нанопорошки железа / Л. В. Коваленко, Г. Э. Фолманис. – М.: Наука, 2006. – 124с. 44. Ковда, В.А. Типы почв, их география и использование / В. А. Ковда, Б. Г. Розанова. – М.: Высшая школа, 1988. – Ч.II. – С.367-378. 45.Кузнецов, В.В. Пролин при стрессе: биологическая роль, метаболизм, регуляция / В. В. Кузнецов, Н. И. Шевякова // Физиология растений. – 1999. – Т.46. – № 2. – С. 321-336. 46. Кривова, Н.А. Состояние слизистой оболочки желудка, про- и антиоксидантной активности и биохимических показателей крови у крыс после скармливания нано- и микрочастиц диоксида титана / Н. А. Кривова, О. Б. Заева, М. Ю. Ходанович и др. // Вестник Томского государственного университета. Биология. – 2011. – №2 (14). – С.81-95. 47. Крылов, С.В. Состав для обеззараживания семян / С. В. Крылов и др. – Патент РФ №2074616 от 10.03.1997 г. 48. Крылов, С.В. Способ предпосевной обработки семян / С. В. Крылов. – Авторское свидетельство №224940 от 3.07.1968 г. 49. Крылов, С.В. Способ оценки метода протравливания семян / С. В. Крылов., Л. А. Паничкин, А. А. Захарин и др. – Патент РФ №2015631 от 15.07.1994 г. 50. Кудоярова, Г.Р. Гидравлическая проводимость корней при гетерогенном распределении элементов минерального питания / Г. Р. Кудоярова, В. К. Трапеникова, И. И. Иванов // Известия Уфимского научного центра РАН. – 2013. – №2. – С. 33-37. 56. Куренева, Е.Н. Использование высокопродуктивных металлов в составе премиксов комбикормов для бройлеров / Е. Н. Куренева, И. А. Егоров, Ю. И. 112 Федоров, Н. Н. Глущенко // Новое в кормлении и содержании сельскохозяйственной птицы. – Загорск, 1984. – С. 3-8. 57. Кушниренко, М.Д. Физиология водообмена и засухоустойчивости плодовых растений / М. Д. Кушниренко. – Кишинев, 1975. – 320 с. 58. Лакин, Г.Ф.Биометрия / Г. Ф. Лакин. – М. : Высшая школа, 1990. – 352 с. 59. Лукина, Н.В. / Н. В. Лукина, В. В. Никонов // Лесоведение. –1999. –№ 2. – С.57-67. 60.Луцик, А.Д. Лектины в гистохимии/ А. Д. Луцик, У. С. Детюк, М. Д. Луцик и др. – Львов.: ВШ, 1989. – 144с. 61. Методические указания 1.2.2635-10 // Медико – биологическая оценка безопасности наноматериалов. – М.: Федеральный центр гигиены и эпидемиологии Роспотребнадзора. – 2010. – 123 с. 62. Мирошникова, Е.П. Обмен химических элементов в организме карпа при использовании наночастиц кобальта и железа в корме / Е. П. Мирошникова, А. Е. Аринжанов, Н. Н. Глущенко, С. П. Василевская и др. // Вестник Оренбургского государственного университета. – 2012. – №6. – С.55-57. 63.Мирошникова, Е.П. Изменение гематологических параметров карпа под влиянием наночастиц металлов / Е. П. Мирошникова, А. Е. Аринжанов, Ю. В. Килякова // Достижения науки и техники АПК. – 2013. – №5. – С. 55-57. 64.Мохаммед, А.М., Ралдугина, Г.Н., Холодова, В.П., Кузнецов, В.В. Аккумуляция осмолитов растениями различных генотипов рапса при хлоридном засолении / А. М. Мохаммед, Г. Н. Ралдугина, В. П. Холодова, В. В. Кузнецов // Физиология растений. – 2006. – Т.53. – № 5. – С.732-738. 65. Нанотехнологии в электронике / под ред. Ю.А. Чаплыгина – М.: Техносфера, 2005. – 448 с. 66. Некрасов, Б.В. Курс общей химии. – М.: Госхимиздат, 1962. – 976 с. 67. Нанотехнология в ближайшем десятилетии. Прогноз направления исследований / под ред. М.К. Роко, Р.С. Уильямса и П. Аливисатоса. М.: Мир, 2002. – 292 с. 113 68. Онищенко, Г.Г. Организация надзора за оборотом наноматериалов, представляющих потенциальную опасность для здоровья человека / Г. Г. Онищенко // Гигиена и санитария. – 2011. – №13. – С. 164-173. 69. Паничкин, Л.А., Райкова, А.П. Использование нанопорошков металлов для предпосевной обработки семян сельскохозяйственных культур / Л. А. Паничкин, А. П. Райкова // Известия ТСХА. – 2009. – Вып.1. – С.59-65. 70. Пассика, К.А. Исследование влияния выбрасов цементной пыли на рост и развитие растений / К. А. Пассика // Успехи современного естествознания. – 2004. – №11. – С. 45. 71. Пахомова, В.М. Основные положения современной теории стресса и неспецифический адаптационный синдром у растений / В. М. Пахомова // Цитология. – 1995. – Т. 37, Вып. 1-2. – С. 66–87. 72. Подколодная, О.А. Пути поступления наночастиц в организм млекопитающих, их биосовместимость и клеточные эффекты/ О. А. Подколодная, Е. В. Игнатьева, Н. Л. Подколодный, Н. А. Колчанов // Успехи современной биологии. – 2012. – №1. – С. 3-15. 73. Поляков, А.Ю. Синтез биосовместимых магнитных наночастиц с различной морфологией и их стабилизация гуминовыми кислотами / А. Ю. Поляков, А. Е. Гольдт, Т. А. Соркина и др. // Перспективные материалы. – 2010. – № 2. – С. 4-16. 74. Пул, Ч., Оуэнс, Ф. Нанотехнологии / Ч. Пул, Ф. Оуэнс. – М.: Техносфера, 2006. – 336с. 75. Райкова, А.П. Предпосевная обработка семян / А. П. Райкова // Материалы международной научно-практической конференции «Льняной комплекс России, проблемы и перспективы. – Вологда, 2001. – С. 9-21. 76. Райкова, А.П. Использование ультрадисперсных порошков металлов для предпосевной обработки семян / А. П. Райкова, Л. А. Паничкин, Н. И. Райкова // Доклады ТСХА. - 2004. – Вып. 276. – С. 44-48. 77. Райкова, А.П. Исследования влияния ультрадисперсных порошков металлов, полученные различными способами на рост и развитие растений / А. П. 114 Райкова, Л. А. Паничкин, Н. Н. Райкова // Материалы международной науч.-техн. конф. «Нанотехнологии и информационные технологии – технологии 21 века». – М., 2006. – С. 118-123. 78. Рахметова, А.А. Ранозаживляющие свойства наночастиц меди в зависимости от их физико-химических характеристик / А. А. Рахметова, Т. П. Алексеева, О. А. Богословская и др. // Российские нанотехнологии. – 2010. – Т.5. – №3-4. – С.102-118. 79. Сабинин, Д.А. Избранные труды по минеральному питанию / Д. А. Сабинин. – М.: Наука, 1971. – 512 с. 80. Селиванов, В.Н. Пролонгированное воздействие ультрадисперсных порошков металлов на семена злаковых культур / В. Н. Селиванов, Е. В. Зорин, Е. Н. Сидорова и др. // Перспективные материалы. – 2001. – №4. – С. 66-69. 81. Серегин, И. В. Физиологические аспекты токсического действия кадмия и свинца на высшие растения / И. В. Серегин, В. Б. Иванов // Физиология растений. – 2001. – Т. 48. – № 4. – С. 606–630. 82. Сизова, Е. А. Наночастицы меди - модуляторы апоптоза и структурных изменений в некоторых органах / Е. А. Сизова, С. А. Мирошников, В. С. Полякова, С. В. Лебедев, Н. Н. Глущенко // Морфология. – 2013. – № 4. –С. 47-52. 83. Спирин, М.Г. Использование обратных мицелл в получении наночастиц золота ультрамалого размера / М. Г. Сирин, С. Б. Бричкин, В. Ф. Разумов // Российские нанотехнологии. – 2006. – Т. 1. – № 1-2. – С. 121-126. 84. Строгонов, Б.П. Физиологические основы солеустойчивости растений / Б. П. Строгонов. – М.: Изд-во АН СССР, 1962. – 366 с. 85. Строгонов, Б.П. Проблемы солеустойчивости растений / Б. П. Строгонов, Л. К. Клышев, Р. А. Азимов и др. – Ташкент: ФАН, 1989. –184 с. 86. Суздалев, И.П. Нанотехнология: физикохимия нанокластеров, наноструктур и наноматериалов / И. П. Суздалев. – М.:КомКнига, 2006. –592с. 87. Сушилина, М.М. Влияние ультрадисперстных порошков металлов (УДПМ) – новых микроудобрений на урожайность и качество зеленой массы 115 :автореф. дис. канд. биол. наук: 06.01.2004 / Сушилина Мария Михайловна. – М., 2004. – 25 с. 88. Сушилина, М.М. Нанотехнологии в растениеводстве и сельском хозяйстве / М. М. Сушилина, А. И. Монькина // Вестник ФГБОУ ВПО РГАТУ. – 2011. – № 3(11). – С.42-44. 89. Титов, А. Ф. Устойчивость растений к тяжелым металлам / А. Ф. Титов, В. В. Таланова, Н. М. Казнина и др. – Петрозаводск: Карельский НЦ РАН, 2007. – 170 с. 90. Трапезников, В.К. Локальное питание растений / В. К. Трапезников, И. И. Иванова, Н. Г. Тельвинская. – Уфа: Гилем, 1999. – 260 с. 91. Удовенко, Г.В. Принципы различной реакции сортов и видов растений на засоление почвы / Г. В. Удовенко // Сорт и удобрение. Иркутск, 1974. – С. 219223. 92. Феник, С.И. Механизмы формирования устойчивости растений к тяжелым металлам / С. И. Феник, Т. Б. Трофимяк, Я. Б. Блюм // Успехи современной биологии. - 1995. – Т. 115. – № 3. – С. 261-275. 93. Фолманис, Г.Э. Ультрадисперстные металлы в сельском хозяйстве / Г. Э. Фолманис. – М., 1999. – 89 с. 94. Франко, О.Л. Осмопротекторы: ответ растений на осмотический стресс / О. Л. Франко, Ф. Р. Мело // Физиология растений. – 2000. – Т.47. – № 1. – С.152159. 95. Хвостов, Д.С. Морфоанатомические и физиологические механизмы приспособления растений земляники к сезонным изменениям условий внешней среды / Д. С. Хвостов // Сельскохозяйственная биология. – 2004. – №5. – С.89-93. 96. Черкозьянова, А.В. Координация роста побега и корня проростков пшеницы в условиях дефицита минерального питания: автореф. дис. канд. биол. наук: 03.00.12 / Черкозьянова Алла Владимировна. – Уфа, 2005. – 139 с. 97. Черняев, Н.Г. Изучение влияния препаратов и способов обработки семян на перезимовку озимых зерновых культур / Н. Г. Черняев // Известие ТСХА. – 1997. – Вып.2. – С.23-28. 116 98. Чижикова, Н.П. Минералы разных гранулометрических фракций как источники элементов питания растений (на примере агросерных тяжелосуглинистых почв Владимирского оползя) / Н. П. Чижикова, Д. В. Карпова // Бюллетень Почвенного Института им. В.В.Докучаева. – 2009. – № 63. – С.41-49. 99. Чиркова, Т.В. Клеточные мембраны и устойчивость растений к стрессовым воздействиям / Т. В. Чиркова// Соросовский образовательный журнал. – 1997. – № 9. – С. 12–17. 100. Чиркова, Т.В. Физиологические основы устойчивости растений / Т. В. Чиркова. – СПб.: Изд. Санкт- Петербургского ун-та, 2002. – 224 с. 101. Шахов, А. А. Солеустойчивость растений / А. А. Шахов. – М. : Изд. АН СССР, 1956. – 552 с. 102. Шевякова, Н.И. Метаболизм и физиологическая роль пролина в растениях при водном и солевом стрессе / Н. И. Шевякова // Физиология растений. – 1983. – Т.30. – № 4. – С.768-783. 103. Шевякова, Н.И. К вопросу о механизмах ответных реакций на засоление различных по солеустойчивости сортов фасоли / Н. И. Шевякова, П. Ковалевски // Сельскохозяйственная биология. – 1994. – № 1. – С.84-88. 104. Юшкин, Н. П. Мир наноминералогии / Н. П. Юшкин, А. М. Асхабов // Вестник института геологии Коми научного центра Уральского отделения РАН. – 2007. – № 12. – С. 2—5. 105. Adili, A. Differential cytotoxicity exhibited by silica nanowires and nanoparticles / A. Adili, S. Crowe, M.F. Beaux // Nanotoxicology. – 2008. – Vol. 2. – Iss. 1. – P. 1 – 8. 106. Allsopp, M. Nanotechnologies and nanomaterials in electrical and electronic goods: A review of uses and health concerns / M. Allsopp, A. Walters, D. Santino // Greenpeace research laboratories. – 2007. – 22 p. 107. Alt, V. An in vitro assessment of the antibacterial properties and cytotoxicity of nanoparticulate silver bone cement / V. Alt, T. Bechert, P. Steinrücke, M. // Biomaterials. – 2004. – Vol. 25. – Iss. 18. – P. 4383-4391. 117 108. Amorim, R. Structures loading with an anticancer compound as drug delivery systems / R. Amorim // Vournal of physical chemistry. – 2012. – V.116 – P. 25642 – 25650. 109. AshaRani, P.V. / P.V. AshaRani, G. Low KahMun // Biomaterials. - 2009.- V. 3.- № 2. P. 279. 110. Atha, D.H. Copper oxide nanoparticle mediated DNA damage in terrestrial plant models / D.H. Atha, H. Wang, E.J. Petersen // Environ. Sci. Technol. – 2012. Vol. 46. - P. 1819−1827. 111. Babu, K. Effect of nano-silver on cell division and mitotic chromosomes / K. Babu, M. Deepa, S. Shankar, S. Rai // The Internet Journal of Nanotechnology. - 2008. Vol. 2. - P. 1−14. 112. Baker, A. J. M. Ecophysiology of metal uptake by tolerant plants / A. J. M. Baker, P. M. Walker // Heavy metal tolerance in plants: evolutionary aspects. Boca Raton: CRC Press. -2008. -Vol. 2. - P. 278. 113. Baun, A. Toxicity and bioaccumulation of xenobiotic organic compounds in the presence of aqueous suspensions of aggregates of nano-C60 / A. Baun, S.N. Sorensen, R.F. Rasmussen // Aquatic Toxicology. - 2008. - Vol. 86. - Iss. 3. - P. 379387. 114. Bogaticov, O.A. Anorganicis nanoparticles in natura / O.A. Bogaticov // Bulletin de RAS. – 2003. – V.73. – n.5 - P. 426-428. 115. Chang, J.Sh. In Vitro cytotoxicitiy of silica nanoparticles at high concentrations strongly depends on the metabolic activity type of the cell line / J.Sh. Chang, K. Liang, B. Chang, D. Hwang, Z. Kong // Environmental Science of Technologies. - 2007. - Vol. 41. Iss. 6. - P. 2064–2068. 116. Chen, L. Manufactured aluminum oxide nanoparticles decrease expression of tight junction proteins in brain vasculature / L. Chen // Journal Neuroimmune Pharmacology. - 2008. - Vol. 3. - Iss.4. - P. 286-295. 117. Chen, Z. Acute toxicological effects of copper nanoparticles in vivo / Z. Chen // Toxicology Letters. - 2006. - Vol. 163.- Iss. 2. - P. 109-120. 118 118. Clemens, S. Toxic metal accumulation, responses to exposure and mechanisms of tolerance in plants / S. Clemens // Biochimie. - 2006. - Vol. 88. - P. 1707–1719. 119. Dachek, W. V. Isolation, assay, biosynthesis, translocation, and function of proline in plant cells and tissues / W.V. Dachek , S. Erickson, S. Sharon // The Botanical Review. - 1981. - V.47. - №3. - P. 349-381. 120. Donaldson, K., Aitken R., Tran L., Stone V., Duffin R. Carbon nanotubes: review of their properties in relation to pulmonary toxicology and workplace safety / K. Donaldson, R. Aitken, L. Tran, V. Stone, R. Duffin // Toxicological Science. - 2006. Vol. 92. - Iss. 1. - P. 5-22. 121. Drexler, K.E. Engines of creation. The Coming Era of Nanotechnology / Drexler K.E. // USA, 1986. – 299 p. 122. Evens, H.J. Potassium and its role in enzyme activation / H.J. Evens, R.A.Wildes // Potassium in Biochemistry and Physiology. - Proc.8. - Int. Potash Inst., Bern. 1971. - P. 13-39. 123. Faisal, M. PhytotoxichazardsofNiO-nanoparticlesintomato: A study on mechanism of cell death / M.Faisal, Q.Saquib, A. A.Alatar // J. Hazard. Mater. – 2013. – Vol. 250-251. – P. 318-332. 124. Geys, J. Acute toxicity and prothrombotic effects of quantum dots: impact of surface charge / J. Geys, A. Nemmar, E. Verbeken // Environmental Health Perspectives. - 2008. - Vol. 116. - № 12. - P. 1607-1613. 125. Ghosh, M. Genotoxicity of titanium dioxide (TiO2) nanoparticles at two trophic levels: plant and human lymphocytes / M. Ghosh, M. Bandyopadhyay, A. Mukherjee // Chemosphere. - 2010. - Vol. 8. - P. 1253–1262. 126. Glass, A.D. Regulation of ion transport / A.D. Glass // Annu. Rev. Plant Physiol. - 1983. - V.34. - P. 311-326. 127. Glass, A.D. The regulation of nitrate and ammonium transport systems in plants / A.D. Glass, D.T. Britto, B.N. Kaiser, J.R. // Journal of Experimental Botany. 2002. -V.53. - P. 855-864. 119 128. Gottschalk, F. Modeled environmental concentrations of engineering nanoparticles (TiO2, ZnO, Ag, CNT, fullerenes) for different regions / F. Gottschalk, T. Sonderer, R.W. Scholz // Environ. Sci. Technol. - 2009. - Vol.43. - P. 9216—9222. 129. Greenway, H. Mechanisms of salt tolerance in nonhalophytes / H. Greenway // Annual Review of Plant Physiology. -1980. - V.31. - P.149-190. 130. Hall, J. L. Transition metal transporters in plants / J. L. Hall, L. E. Williams // Journal of experimental botany. - 2003. - Vol. 54. - P. 2601–2613. 131. Haschke, K.P., Luttge K. Interactions between IAA, potassium and malate accumulation and growth in avenacoleopite segments / K.P. Haschke, K. Luttge // Pflanzenphysiol. – 1975. - V.76. - P. 450-455. 132. Heinlaan, M. Toxicity of nanosized and bulk ZnO, CuO and TiO2 to bacteria vibrio fischeriand crustaceans daphnia magna and thamnocephalusplatyurus / M. Heinlaan, A. Ivask, I. Blinov // Chemosphere. - 2008. - Vol. 71. - P. 1308-1316. 133. Hillegass, J. M. Interdiscip.Rev.Nanomed / J. M. Hillegass, A. Shukla, , S. A. Lathrop // Nanobiotechnol. - 2010. - V.2. - P. 219-231. 134. Hong, S. Interaction of Poly(amidoamine) Dendrimers with Supported Lipid Bilayers and Cells: Hole Formation and the Relation to Transport / Hong S., Bielinska A.U., Mecke A. // Bioconjugate Chemistry. - 2004. - Vol. 15. - P. 774–782. 135. Ji, J.H. Twenty-eight-day inhalation toxicity study of silver nanoparticles in Sprague-Dawley rats / J.H. Ji // Inhalation Toxicology. - 2007. - Vol. 19. P. 857-71. 136. Jiang, J. Does nanoparticle activity depend upon size and crystal phase / J. Jiang, G. Oberdrster, A. Elder, R. Gelein // Nanotoxicology. - 2008. - Vol. 2. - P. 138146. 137. Kahru, A. Biotests and biosensors for ecotoxicology of metal oxide nanoparticles: a minireview / A. Kahru, H. Dubourguier, I. Blinova, A. Ivask, K. Kasemets // Sensors. - 2008. - Vol. 8. P. 5153 - 5170. 138. Kang, S.J. Titanium dioxide nanoparticles trigger p53-mediated damage response in peripheral blood lymphocytes / S.J. Kang // Environmental Molecules Mutagens. - 2008. - Vol. 49.- P. 399-405. 120 139. Kaura I.P., Bhanarib R., Bhanarib S., Kakkara V. Potential of solid lipid nanoparticles in brain targeting // J. of Controlled Release. - 2008. - Vol. 127. - Iss. 2. P. 97-109. 140. Klancnik, K. Use of a modified Allium test with nanoTiO2 / K. Klancnik, D. Drobne // Ecotoxicol. Environ. Saf. - 2011. - Vol. 74. - P. 85 - 92. 141. Krämer, U. Transition metal transport / U. Krämer, I.N. Talke, M. Hanikene // FEBS Letters. - 2007. - Vol. 581. - P. 2263 - 2272. 142. Kravchyshyn, M.D. Materiales composition agueum suspension Northern Dvinaaestuario (albus mare) dumvernumaestus / M.D. Kravchyshyn M.D. // Oceanology. – 2010. - P. 396 - 416. 143. Lead, J.R. Natural aquatic colloids: current knowledge and future trends / J.R. Lead, K.J. Wilkinson // Environ. Chem. - 2006. - Vol. 3. - P. 159-171. 144. Lee, C.W. Developmental phytotoxicity of metal oxide nanoparticles to Arabidopsis thaliana / C.W. Lee, S. Mahendra, K. Zodrow // Environ. Toxicol. Chem. 2010. - Vol. 29. - P. 669-675. 145. Lee, W. M. Toxicity and bioavailability of copper nanoparticles to the terrestrial plants mung bean (Phaseolusradiatus) and wheat (Triticumaestivum): plant agar test for water-insoluble nanoparticles / W. M. Lee, Y. J. An, H. Yoon, H.S. Kweon // Environ. Toxicol. - 2008. - Vol. 27.- P. 1915−1921. 146. Lei, R., Wang, Q. // Toxicol. - 2008.- V.232. - P. 292-301. 147. Leigh, R.A. An hypothesis relating critical potassium concentrations for growth to the distribution and functions of this ion in the plant cell // New Phytologist. 1984. - V.97. - P.1-13. 148. Lesniak, W. Dendrimernanocomposites as biomarkers: fabrication, characterization, in vitro toxicity and intracellular detection / W. Lesniak, A. Bielinska, K. Sun // Nanoletters. - 2005. - Vol. 5. - P. 2123-2130. 149. Lewinski N. Cytotoxicity of Nanoparticles / Lewinski N., Colvin V., Drezek R. // Small-journal. - 2008. - №1. - Р. 26 – 49. 150. Lin, D. Phytotoxicity of nanoparticles: inhibition of seed germination and root growth / D. Lin // Environmental Pollutants. - 2007. - Vol. 150. Iss. 2. - P. 243-250. 121 151. Lu, N. Nano titanium dioxide photocatalytic protein tyrosine nitration: a potential hazard of TiO2 on skin / N. Lu // BiochemBiophys Res Commun. - 2008. Vol. 370. - Iss. 4. - P. 675-680. 152. Mahmoudi, M. Assessing the in vitro and in vivo toxicity of superparamagnetic iron oxide nanoparticles / M. Mahmoudi, H. Hofmann // Chem. Rev. - 2012. - Vol.112(4). - P.2323 - 2338. 153. Markovic, Z. The mechanism of cell-damaging reactive oxygen generation by colloidal fullerenes / Z. Markovic, B. Todorovic-Markovic, D. Kleut, N. Nikolic // Biomaterials. - 2007. - Vol. 28. - P. 5437-5448. 154. Mehrle, P.M. Biomarkers: Biochemical, Physiological, Histological Markers of Anthropogenic Stress / P.M. Mehrle, H.L. Bergmann // USA, 2002. – Р. 211–234. 155. Musante, C., White J.C. Toxicity of silver and copper to Cucurbitapepo: differential effects of nano and bulk-size particles / C. Musante , J.C White // Environ. Toxicol. - 2012. - Vol. 27. - P. 510 - 517. 156. Nel, A.E. Understanding biophysicochemical interactions at the nano-bio interface / A.E. Nel, L. Madler, D. Velegol // Nat. Mater. - 2009. - Vol.8. - P. 543—557. 157. Ostiguy, C. Health effects of nanoparticles / C. Ostiguy, G. Lapointe, M. Trottier // Studies and research projects. - 2006. - P.52. 158. Prasad, M. N. Cadmium toxicity and tolerancein vascular plants / M. N. Prasad // Environ. Exp. Bot. - 1995. - Vol. 35.- P. 525–545. 159. Price, N.M. Selenium: An essential element for growth of the coastal marine diatom / N.M. Price // J. Phyol. - V.23. -P. 1-9. 160. Prow, T. Construction, gene delivery, and expression of DNA tethered nanoparticles / T. Prow, J.N. Smith, R. Grebe // Molecular Vision. - 2006. - Vol. 12. - P. 606-615. 161. Reevesa, J.F. Hydroxyl radicals (OH) are associated with titanium dioxide (TiO2) nanoparticle-induced cytotoxicity and oxidative DNA damage in fish cells / J.F. Reevesa // Mutation Research / Fundamental and Molecular Mechanisms of Mutagenesis. - 2008. - Vol. 640. Iss. 1-2.- P. 113-122. 122 162. Satter, R.L. Rhythmic potassium flux in Allbizzia. Effect of aminophylline, cations and inhibitors of respiration and protein synthesis / R.L. Satter, P.B. Applewhite, A.W. Galston // Plant Physiology. - V.54. - P. 280-285. 163. Schranda, A.M. Differential biocompatibility of carbon nanotubes and nanodiamonds / A.M. Schranda, L. Daia, J.J. Schlager, S.M. Hussain, E. Osawa // Diamond and Related Materials. - 2007. - Vol. 16. - P. 2118-2123. 164. Singer, P. Nanotechnology / P. Singer // Semiconductor International. 2007. - P. 36 - 40. 165. Soenen, S.J. Himmelreich U., Nuytten N., De Cuyper M. Cytotoxic effects of iron oxide nanoparticles and implications for safety in cell labeling / S.J. Soenen , U. Himmelreich , N. Nuytten , M. De Cuyper // Biomaterials. - 2011. - Vol. 32(1). - P. 195 - 205. 166. Stampoulis, D. Assay-dependent phytotoxicity of nanoparticles to plants / D. Stampoulis, S. K. Sinha, J.C. White // Environ. Sci. Technol. - 2009. - Vol. 43. - P. 9473 - 9479. 167. Stewart, G.R. The rate of Proline Accumulation in Halophytes / G.R. Stewart, J. A. Lee // Planta Berl. - 1974. - Vol. 120. - P. 279-289. 168. Veen, B.W. The role of nitrate in osmoregulation of Italian ryegrass / B.W. Veen, A. Kleinendorst // Plant and Soil. – 1986. – V.91. – P.433-436. 169. Wagner, G. J. Accumulation of cadmium in crop plants and consequences to human health / G. J. Wagner // Advances in Agronomy. – 1993. – Vol. 51. – P. 173– 212. 170. Wang, B. Acute toxicity of nano- and micro-scale zinc powder in healthy adult mice / B. Wang // Toxicology Letters. – 2006. – Vol. 161. – Iss. 2. – P. 115-123. 171. Werner, D. Silica metabolism.In: Inorganic Plant Nutrition. Encyclopedia of Plant Physiology, New Series. Eds. A. Lauchli, R.L. Bieleski / D. Werner, R. Roth // Berlin: Spring-Verlag, 1983. – V.15B. – P.682-694. 172. Wu, W.H. Rare earth elements activate endocytosis in plant cells / W.H. Wu, X. Sun // Biochem. Biophys. Res. Commun. – 2008. – V. 373. – №2. – P. 315. 123 173. Yushkin, N.P. Mineralis mundi et biosphere: mineralisorganiz-mobioz, biomineral interaction coevolution / N.P. Yushkin // Acta iv International seminar «Mineralogy et vitam: Origin de biosphere et co-evolution of mineralis et biologicummundosbiomineral-ogiya» (Syrtyvkar, Komi Republie, May 22-25, 2007). Syrtyvkar, 2007 – P. 5-7. 174. Zhu, M.T. Comparative study of pulmonary responses to nano- and submicron-sized ferric oxide in rats / M.T. Zhu, W.Y. Feng, B. Wang // Toxicology, 2008. – Vol. 247. – Iss. 2-3. – P. 102 - 111. 175. Zhua, S. Toxicity of an engineered nanoparticle (fullerene, C60) in two aquatic species, Daphnia and fathead minnow / S. Zhua, E. Oberdörsterb, M.L. Haascha // Marine Environmental Research. – 2006. – Vol. 62. – P. 5-9.