ВОЗМОЖНЫЙ ОЛИГОНУКЛЕОТИД-ЗАВИСИМЫЙ МЕХАНИЗМ АДАПТАЦИЙ Дейчман А.М.
реклама

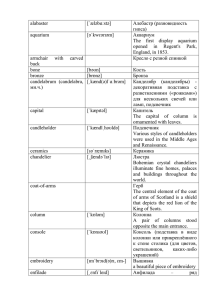
ВОЗМОЖНЫЙ ОЛИГОНУКЛЕОТИД-ЗАВИСИМЫЙ МЕХАНИЗМ АДАПТАЦИЙ Дейчман А.М. ФГБУ Российский Онкологический Научный Центр им. Н.Н.Блохина, Российской Академии Медицинских Наук, e-mail : amdeich@rambler.ru Проблема молекулярных, клеточных и, далее, системных механизмов онтогенетических и филогенетических адаптаций исходно тесно связана с проблемами закрепления дифференцировочных состояний клетки (структуры ее генома), – с участием известных / неизвестных генетических и эпигенетических факторов и механизмов регуляции и наследования. Одним из до сих пор не показанных (вероятно фундаментальных) механизмов, связанных с в.н. регуляцией / наследованием, может оказаться гипотетический механизм воспроизведения клеткой условно коротких (15-30 нуклеотидов) и средних [(1530)n≤5] олигонуклеотидных последовательностей, способных, одновременно, к выполнению широкого круга реактивно-регуляторных (включая защитные для клетки/организма) и более длительно действующих, эволюционно-значимых нуклеотидно-пластических функций в геноме. Воспроизведение олигонуклеотидов было названо вПОТ-механизмом, – вариабельной Поэпитопной Обратной Трансляции отдельного эпитопа (в 5-10 аминокислот). В этом случае, концевые аминокислоты эпитопов, локализованных на внутренних мембранах митохондрий (труднопроницаемых даже для малых ионов), последовательно вырезались протеиназами и связывались с соответствующими тРНК (или ориентировали вблизи себя наборы Аа-тРНК). Далее, сближенные антикодоновые участки тРНК (АатРНК): 1. либо выполняли роль псевдоматрицы для полимеразной активности, прерывисто синтезирующей рибо- / дезоксирибо-олигонуклеотиды (и, возможно, их комплементарные копии); 2. либо последовательно вырезались и лигировались. В обоих случаях, в силу разных причин, могло формироваться несколько вариантов нуклеинового эквивиалента (НЭ в 15-30 нуклеотидов; новая «минимальная генетическая единица») данного эпитопа, отсюда – вариабельность вПОТ-механизма (у прокариот подобный механизм может быть ассоциирован с цитоплазматической мембраной; в разработке). Перенос НЭ (митохондрии→ядро / цитоплазма) может быть связан с транспортирующими их векторными (ретровирус-подобными) нуклеиновыми последовательностями (ВНП-передача) и белками РНП / ДНП-комплесов. Такие НЭ могут частично/полностью-комплементарно взаимодействовать: 1) с короткими РНК (микроРНК, малыми интерферирующими РНК, др.; всего около 10 видов) и их мРНК / РНК-мишенями (к которым, в частности, относятся белки, модифицирующие гистоны, негистоновые белки, метилирующие ДНК, т.е. формирующие т.н. «гистоновый код», конформационную структуру нуклеосом и «метилом» ДНК); 2) с РНКкомпонентами РНП-комплкексов в цитоплазме / ядре (влияют на трансляцию, экспрессию генома); 3) с ориджнами репликации (ядра / митохондрий / хлоропластов), 4) с сайтами инициации транскрипции, – и 5) с праймерами тех и других; а также с сайтами: 6) «горячих точек» генома, 7) репарации одно- / двунитевых разрывов, 8) сплайсинга, 9) повторяющихся последовательностей (вблизи генов, подвижных элементов). Таким образом, НЭ может позитивно / негативно вмешиваться в такие процессы регуляции экспрессии генома, как репликация, транскрипция, трансляция, сплайсинг, генный / геномный сайленсинг, редактирование РНК, др. Кроме того, единичные (15-30)- и амплифицированные (15-30)n-варианты НЭ могут служить тем исходным материалом, из которого, за эволюционно значимый период, формируются целые геномы организмов: прежде всего повторы (простые тандемные, др.; найдены в межгенных, некодирующих, нетранслируемых, интронных, промоторных, экзонных, др. регуляторных областях генома эукариот), из которых, далее, воспроизводится большая часть генома, а также интроны и экзоны. Именно пунктуация повторов задает рамки и предпочтительные направления, в которых осуществляется генетический метаболизм любых организмов (различные виды рекомбинации). Повторы, не редко, имеют сайты гипервариабельности / нестабильности; именно они являются наиболее полиморфными областями генома. С повторами связывают многие генетические заболевания, а также раки (прямой кишки, легких, молочной железы), при которых обнаруживают MSI-сайты микросателлитной нестабильности. Напряженно функционирующий вПОТ-механизм (генетического поиска) обоюдоостр, и может вести к генетической нестабильности и патологическим результатам за счет внедрения векторов (с НЭ внутри) в перечисленные участки генома. В процессе экспрессии генома, предложенный вПОТ-механизм, как и синтез белка, используется большинством клеток (соматических, половых, стволовых, иммунокомпетентных, др.) любых организмов в соотвествии с их генетической программой. Связанные с вПОТ / ВНП-передача механизмами изменения регуляции и нуклеотидного состава генома соматических клеток (в течение жизни организма) могут вести к изменению регуляции генных-сетей, генетической/эпигенетической регуляции, и, в частности, к онтогенетическим адаптациям (возможны и соматические нейтральные / летальные / апоптогенные / др. мутации). В соответствии с воззрениями Ч.Дарвина (т.н. «теорией пангенезиса»), не знавшего про ДНК/РНК, некоторые из изменений в соматических клетках могут быть переданы (в виде «геммул») в герминативные клетки. Механизм передачи / закрепления новой генетической информации от соматических клеток к половым связан с пороговым преодолением каждого из множества этапов/барьеров этого процесса, потенциально, и с определенной небольшой частой, завершающегося филогенетической адаптацией. Работы автора представлены на сайте (www.amdeich-var-reverse-translation.ru). THE POSSIBLE OLIGONUCLEOTIDE-DEPENDENT MECHANISM OF ADAPTATIONS Deichman A.M. FSBA Russian Cancer Research Center by N.N.Blokhin, Russian Academy of Medical Sciences, Moscow, Russia e-mail: amdeich@rambler.ru The problem of molecular, cellular, and further systemic mechanisms of ontogenetic and phylogenetic adaptations closely linked initially to the problems of fixing differentiated cell states (the structure of its genome), – with the participation of known / unknown genetic and epigenetic factors and mechanisms of regulation and inheritance. One has not yet been shown (probably basic) mechanisms associated with above named regulation/ /inheritance, it may be a hypothetical mechanism of cell reproduction relatively short (15-30 nucleotides) and medium-sized [(15-30)n≤5] oligonucleotide sequences, capable simultaneously to perform a wide range of reactive-regulatory (including protective for the cell / body) and a long-acting evolutionarily significant nucleotideplastic functions in the genome. Reproduction of oligonucleotides was called vIERT-mechanism, – variable Individual Epitope Reverse Translation of single epitope (in 5-10 amino acids). In this case, the terminal amino acids of epitopes localized on the inner membranes of mitochondria (poorly permeable even for small ions), consistently cut by proteases and associated with the corresponding tRNA (or oriented near a set of Aa-tRNA). Furthermore, the contiguous of the tRNA anticodon sites (Aa-tRNA): 1. or acted as pseudomatrix for polymerase activity, discrete synthesizing ribo-/ /deoxyribooligonucleotides (and possibly their complementary copies); 2. or they were sequentially cut and ligated. In both cases, for various reasons, may have formed the several variants of nucleic equivalents (NE in 1530 nucleotides, a new "minimal genetic unit") of the epitope, from here – variability of vIERT-mechanism (in prokaryotes such a mechanism may be associated with the cytoplasmic membrane; in preparation). Transfer of NE (mitochondria → nucleus / cytoplasm) may be associated with transporting them by vector (retrovirus-like) nucleic acid sequences (VLNS-transfer) and proteins of RNP / DNP-complexes. These NEs may partially / fully-complementary interact: 1) with short RNAs (micro RNA, small interfering RNA, etc.; about 10 species) and their mRNA/RNA-targets (proteins which, in particular, modify histones, nonhistone proteins, methylating DNA, i.e., forming the so-called "histone code", the conformational structure of the nucleosomes and "methylom" DNA); 2) with the RNA-components of RNP-complexes in the cytoplasm / nucleus (effect on translation, expression of genome); 3) with origins replication (nuclei / mitochondria / chloroplasts), 4) with sites initiation of transcription, 5) with primers of both; as well as the sites: 6) "hot spots" of the genome, 7) repair single- / double-strand breaks, 8) splicing, 9) repetitive sequences (nearby of genes, mobile elements). Thus NE may have a positive / negative to interfere in these processes regulate the expression of the genome, as replication, transcription, translation, splicing, gene/genomic silencing, RNA editing, etc. In addition, the unit (15-30)- and amplified (15-30)n-versions of NEs can be used as initial material from which for significant evolutionary period formed the entire genomes of organisms: first of all repeats (simple tandem, etc.; are found in intergenic, non-coding, untranslated, intron, promoter, exon, and oth. regulatory regions of the genome of eukaryotes) of which furthermore reproduced a large part of the genome, as well as introns and exons. Exectly punctuation of the sets repeats define the frameworks and the preferred directions in which the genetic metabolism (different types of recombinations) have a place to be for any organism. In repeats are not uncommon sites of hypervariability / instability, they are the most polymorphic regions of the genome. Repeats associated with many genetic diseases and cancers (colorectal, lung, breast) in which sites exhibit MSI-microsatellite instability. Intensive-functioning vIERT-mechanism (genetic search) is double-edged, and can lead to genetic instability and pathological results due to the introduction of vectors (with NE inside) in the above call sites of the genome. In genome expression process suggested vIERT mechanism, as well as protein synthesis, is used by most cells (somatic, germinal, stem, immune cells, etc.) of any organisms in accordance with their genetic program. Related vIERT / VLNS-transfer mechanisms and changes in the regulation of the nucleotide composition of the genome of somatic cells (during the life of the organisms) can lead to a change in the regulation of gene networks, genetic / epigenetic regulation, and in particular to the ontogenetic adaptations (possible and somatic neutral / lethal / apoptogenic / oth. mutations). In accordance with the views of Ch.Darwin (so-called "theory of pangenesis"), who did not know about the DNA / RNA, some of the changes in somatic cells can be transferred (in the form of "gemmules") in the germinal cells. The mechanism of transfer / fixation of new genetic information from somatic cells to germinal cells is associated with overcoming the threshold of each of the many steps / barriers of this process, potentially and with some small occurrence, culminating in a phylogenetic adaptation. The author's work presented at this site (www.amdeich-var-reverse-translation.ru).